4. Relationship between prostephanus truncatus teretriosoma nigrescens
4.1 Influence of teretriosoma nigrescens under near-to-practical conditions
The influenced of the imported useful insect, T. nigrescens on the non-native pest P. truncatus was to be investigated in Togo under quarantine conditions prior to release of the predator. For this, types of maize stores common in southern Togo were erected in a wire gauze cage which prevented immigration and emigration of the insects. However, in order to obtain a spectrum of pests in the stores as would occur under natural conditions, the maize was firstly exposed to natural infestation by pests over a period of 4 months prior to closing the cage and prior to introducing the predator.
4.1.1 Material and methods
In August 1989, 6 test stores of the traditional model "Ebli-va" were erected on a concrete platform (6 x 10 in) which had been prepared (Fig. 17). A platform made of the commonly used types of wood (length 120 cm, width 120 cm) was mounted on the four legs of the wooden supports (height 80 cm) of the stores. The maize (approx. 200 kg of a local, yellow-grained variety per store) was stored on 23rd and 24th August, 1989 On 15th and 16th November 1989, the maize cobs were removed from all stores, systematically mixed and finally restored. This was done to ensure a homogenous content of pests in all stores.
Test cage
The gauze cage (B x L x H: 5 x 9 x 2.50 m) (Fig. 17) was divided into two halves along its length. Four cells behind each other were contained in each half in each case, these were 3 test cells (2.50 x 2.50 m) and a transfer cell (1.50 x 2 so m). There was no connection between the two halves. In each half each cell was connected to the next by a door. One half of the total cage was used for the three repetitions of an experiment. The frame of the cage was made of flat iron treated with rust proofing paint.
The stainless steel gauze (0.5 mm mesh) was stretched between the flat iron and, to ensure it was absolutely sealed, covered in a plastic sealing. Water was drained off through siphons, separated for each half of the cage, which each led into a closed drainage container.
Adding T. nigrescens
The adult specimens of T. nigrescens were transported from the BBA (Federal Biological Research Centre for Agriculture and Forestry) in Berlin to Lom� according to quarantine regulations, on 17th November 1989.
Prior to transport, 5-6 generations of the predator, which was caught in Costa Rica in November 1988, were observed, bred and examined for pathogens under quarantine at the BBA. The insects were free of any pathogens and of any antagonist. 2000 adults of the predator for each repetition were introduced into the quarantine cage on 18th December 1989.
Sampling and evaluation
The first sampling of the maize took place directly after it went mm storage in August 1989. After two further 100-cob samples at intervals of one month, a pause was made during which the cobs were mixed, new stores were erected and the cage was built over the stores. After closing the cage, but prior to introducing T. nigrescens, regular sampling was resumed. The samples were gassed with hydrogen phosphide in a metal box in the transfer cells of the cage for at least a week before they were evaluated. The single cob method for the total 100-cob sample was used for evaluation, and calculation of loss was according to the CWM (Chapter 2.1.2).
Illustration of the results of the investigation at this point is restricted to the most significant primary pests and loss parameters. The results can be seen in full in HELBIG (1993).
4.1.2 Populations of pests and useful insects
The interaction between P. truncatus and T. nigrescens, and the resulting development in the population of the pest forms the central element of this investigation. In addition however, the population development of the other pests is to be noted due to a possible relationship of the predator with these pests.
P. truncatus
An influence of the predator on the population development of the pest became visible 2 months after it was introduced in the number of adult P. truncatus found (Fig. 18). At this early point, the difference between the two experiments was relatively small and not yet significant at 301 beetles. During the following months, strong growth of the pest population could be observed in the experiment without T. nigrescens. This led to a maximum of 4086 adult P. truncatus per 100 cobs after 8 months.
In comparison to the development of the population in the control, the growth of the pest population in the variant with T. nigrescens was very weak. The highest number of adult P. truncatus counted after 7 months and amounted to 1117 beetles. After the 9 months of the experiment, the number of adult P. truncatus amounted to 760 in presence of the predator and was 79.8% lower than in the control containing 3670. T. nigrescens was thus able to very effectively suppress the population development of the Larger Grain Borer. Since only the number of adult insects was counted in this experiment, the effect of the predator did not become clearly visible until after a certain time.
{kind=link}
T. nigrescens
The transport of T. nigrescens from Costa Rica via Berlin to Togo meant a new habitat for the predator. The environmental conditions in this new habitat are different from those in its native home in Central America. Since this experiment was carried out in a closed wire-gauze cage, the number of environmental factors was thus limited. The major factors influencing the predator can thus be seen as the climatic conditions.
T. nigrescens evidently had no serious problems in adapting to the new environmental conditions (Fig. 19). The population increased during the whole course of the experiment and, after 9 months, reached am average density of 876 adult specimens per 100 cobs.
If the abundance of T. nigrescens is related to the population of P. truncatus a prey-predator ratio of 1:1.2 after 9 months is produced. Although a very close prey-predator ratio of 3.6:1 was determined at the beginning of the experiment, in comparison to surveys under practical conditions in Costa Rica (see B�YE, 1988), this ratio became even closer during the course of the experiment.
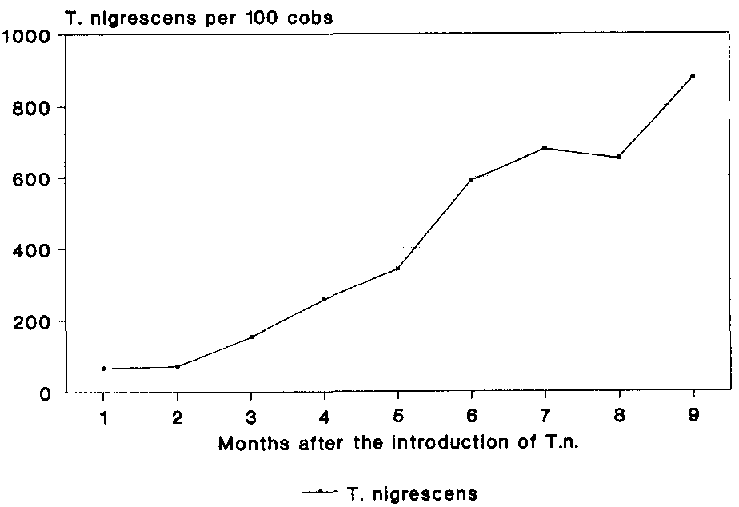{kind=link}
S. zeamais
Apart from P. truncatus, the most important pest responsible for significant damage and losses to stored maize is S. zeamais. It thus becomes interesting to observe the population development of this pest and the structure of the loss parameters. In add hon. a comparison of the development of the population of S. zeamais with and without a predator and under the aspect of possible side-effects of using T. nigrescens is important.
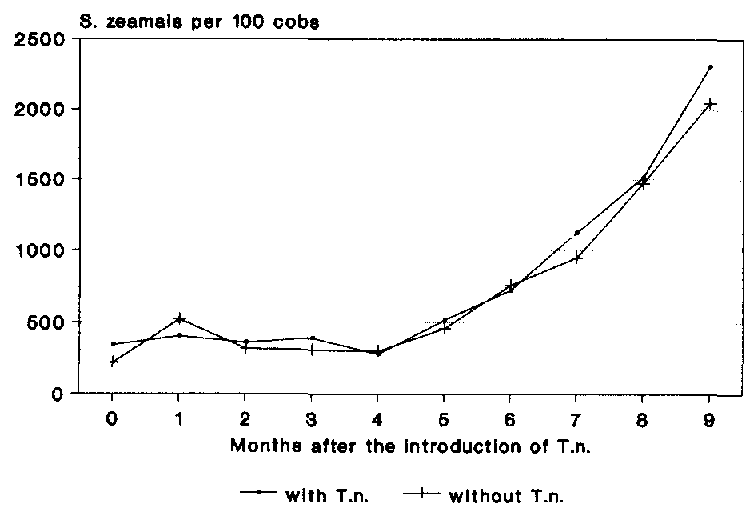
The population density of S. zeamais remained at the initial level of approx. 300400 beetles per cob (Fig. 20) in both experiments up to the fourth month of storage. Growth in the population d d not become visible until the fifth month. During the following months, the populations in both experiments underwent almost parallel development and, after the experimental period of 9 months, reached a density of 2048 and 2308 beetles per 100 cobs respectively. No influence of T. nigrescens could be observed on the population development of S. zeamais in this experiment.
{kind=link}
Secondary pests
The populations of secondary pests also have to be considered regarding any side-effects occurring in the use of T. nigrescens. Whilst observing and evaluating the development of the population, the dependency of secondary on primary pests has to be particularly noted. Especially when maize is stored as cobs, infestation by primary pests is absolutely essential before secondary pests are able to make use of the nutritive substrate.
P. truncatus produces a substantial quantity of boring powder which forms the staple food for secondary pests. This makes their development directly dependent on the boring powder and indirectly dependent on the population of the Larger Grain Borer. The influence of T. nigrescens on the population of P. truncatus, and as a result of this, on quantity of boring powder, thus becomes an indirect effect. In this experiment it is not possible to distinguish any direct from indirect influence due to the methods employed.
As the test stores in the wire-gauze cage were exposed to natural infestation, the secondary pests occurring were the same as in farm-level storage: Carpophilus spp., C. quadricollis, T. castaneum, Cryptolestes spp., P. subdepressus and O. surinamensis. The development of the populations of Carpophilus spp., C. quadricollis and O. surinamensis were almost identical in the stores with and without the predator. For Cryptolestes spp. and T. castaneum slightly reduced density was recorded for the stores containing T. nigrescens, whilst P. subdepressus showed distinctly reduced abundance.
Where Cryptolestes spp. and P. subdepressus are concerned, this is likely to have been an indirect effect of T. nigrescens. In the variant with the predator, the population of the Larger Grain Borer was considerably reduced (see above) and consequently the amount of boring powder produced was smaller. The basic nutritive conditions were thus worse for the secondary pests in this variant. This assumption could also apply to T. castaneum. This pest is also familiar as a predator which could mean that the smaller supply of prey was also the reason for the slight reduction in population density in the variant with the predator.
4.1.3 Damage and loss situation
In addition to recording the development of the population for the pests, the damage and loss situation was analysed. The intention was to find indications of the efficacy of T. nigrescens in relation to the losses caused by P. truncatus. As the experiment was not carried out under practical but near-to-practical conditions, its significance is restricted. Nevertheless, it does provide initial orientation on how the predator possibly contributes to reducing the loss occurring under practical conditions.
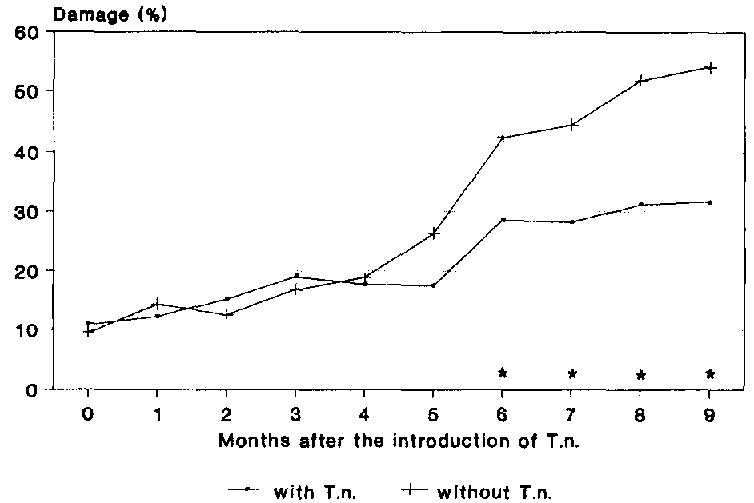
At the time of introducing the predator T. nigrescens to the maize stores, the damage amounted in both variants to approx. 10% (Fig. 21). in the subsequent four months, the amount of damage rose slightly, reaching a value of approx.20% in both experiments. From this point onwards, there was a distinct difference evident between the two variants.
{kind=link}
In the variant without the predator, a rapid increase damage was recorded from the fifth month of storage onwards, which continued until the end of the observation period. After 9 months of storage, the damage in the control amounted to 54.1%.
In the maize stores, to which T. nigrescens had been introduced, an increase in damage was also measured, however this remained considerably less than in the control. With 20% damage in the fifth month of storage, this rose to approx. 30% in the sixth month of storage. Subsequently, damage remained at this level and amounted to 31.6% after the nine months of storage, meaning a statistically significant reduction in comparison to the control of 41 .6%.
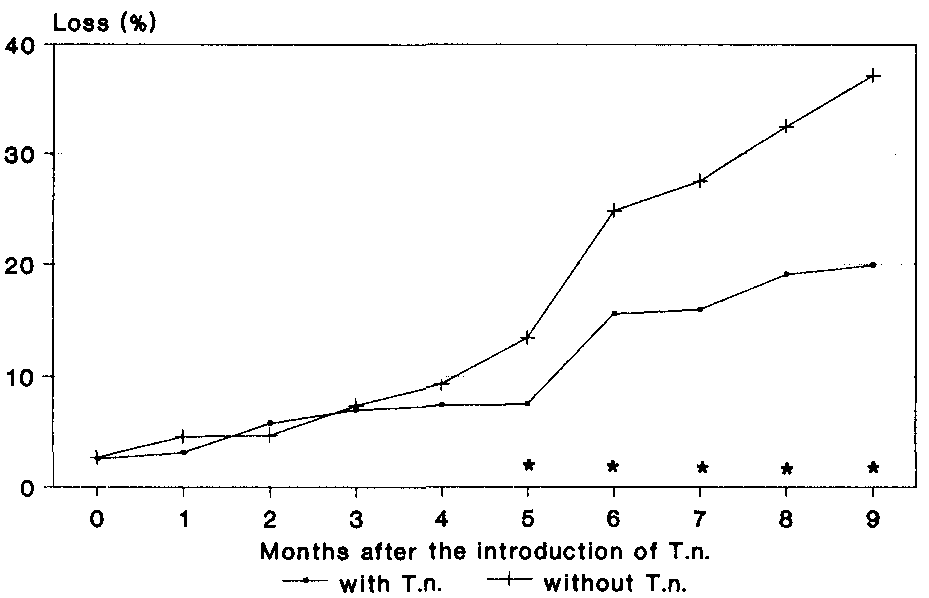
Beginning with 3%, loss within the first three months after the introduction of the predator only increased a little and rose to 7% (Fig. 22). After this point, loss in the maize stores without the predator increased considerably more than in the variant with T. nigrescens. This development continued until the end of the observation period After 9 months of storage, the dry-weight loss was recorded to be 37.2% in the control.
{kind=link}
In the maize stores with the predator, an increase loss was also recorded, but this was lower than in the comparison and decreased as the experimental period progressed. After 9 months of observation, the loss recorded in the presence of T. nigrescens was 19.9% which was 46.5% lower than in the maize stores without the predator.
The influence of T. nigrescens on the structure of damage
An analysis of the damage structure to the maize cobs was also made in this experiment. The structure of damage cam be used as am orientation for the losses of edible maize which the farmers suffer due to infestation by insects (see Chapter 2.6). Allocation into categories followed the key given in Chapter 2.1.2. Maize cobs showing more than 75% damaged grains were allocated to category 5.
In the maize stores without the predator, the proportion of cobs in category 5, with an initial level of 7% at the beginning of the experiment, increased over the whole period of storage (Tab. 18). After 9 months of observation, 56% of the cobs had been allocated to category 5. Assuming that the cobs in category 5 were no longer suitable for consumption, this would mean a loss of edible maize amounting to 56%.
In the stores with the predator, the proportion of cobs in category 5 developed parallel to the control in the first months of storage (Tab. 19). This phenomenon was also observed for other loss parameters. From the fifth month onwards, a difference developed between the two variants with a clearly slower increase in the number of cobs being allocated to category 5 in the presence of the predator.
A striking feature is the fast increase in the percentage allocated to class 5 from the fifth to the sixth month after beginning the experiment. This leap is also reflected in the developments of damage, loss and production of boring powder.
The development in the population of P. truncatus, did not run parallel to this, but the population of S. zeamais showed a strong increase in this period. The damage could thus have been caused by this pest (cf. Chapter 4.1.2). After 9 months of experimentation, 31% of the cobs in the presence of T. nigrescens were in class 5. The loss of edible maize was 44% lower than in the variant without the predator.
Tab18: Structure of the damage to maize cobs in the absence of the predator Teretriosoma nigrescens(proportion (%) of 300 cobs for each point in time in individual categories of damage)
| Cate- gory |
Months after the introduction of the predator | |||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | 61 | 61 | 60 | 54 | 53 | 44 | 36 | 34 | 25 | 27 |
| 2 | 27 | 23 | 24 | 24 | 24 | 15 | 20 | 17 | 14 | |
| 3 | 2 | 1 | 2 | 4 | 4 | 1 | 6 | 3 | 3 | 1 |
| 4 | 3 | 1 | 2 | 0 | 1 | 3 | 3 | 1 | 2 | 2 |
| 5 | 7 | 14 | 11 | 18 | 18 | 28 | 40 | 43 | 53 | 56 |
Tab. 19: Structure of the damage to maize cobs in the presence of the predator Teretriosoma nigrescens (proportion (%) of 300 cobs for each point in time in individual categories of damage)
| Cate- gory |
Month after introduction of the predator | |||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | 50 | 56 | 56 | 54 | 50 | 53 | 46 | 43 | 39 | 39 |
| 2 | 35 | 29 | 25 | 25 | 30 | 26 | 23 | 27 | 28 | 27 |
| 3 | 3 | 2 | 3 | 3 | 1 | 3 | 1 | 2 | 3 | 1 |
| 4 | 4 | 4 | 2 | 2 | 1 | 0 | 1 | 1 | 3 | 2 |
| 5 | 8 | 9 | 14 | 17 | 19 | 18 | 28 | 28 | 27 | 31 |
4.2 Comparison of the efficacy of teretriosoma nigrescens in Togo and Costa Rica
In a comparison of P. truncatus, from different origins, NISSEN (1989) recorded extensive differences in the fitness of the pest. The fitness of the pest population could significantly affect efficacy of a useful insect during biological control. Initial orientation regarding the efficacy of the predator to be expected in Togo, as compared to in the native home of the predator and the useful insect is to be provided by this experiment. For this, the conditions under which the experiment was carried out were taken from the work of B�YE (1988) carried out in Costa Rica. Ensuring conditions as similar as possible to this makes a direct comparison of the results possible
Material and methods
20 adult P. truncatus, were put onto 200 g shelled maize of a local white-grained variety. After 7 days, T. nigrescens were added on a ratio (P.t.: T.n.) of 2:1 and 3.3:1. The experiment lasted 50, 70 and 110 days. The experiments were carried out in 11 glass vessels in the transfer cell of the wire-gauze cage at a temperature of 19-38 �C and a relative humidity of 25-99%. Evaluation was carried out according to the method described in Chapter 2.1.2.
Results
The development of the pest population was clearly reduced in the presence of the predator, independent of the initial ratio of individuals (Tab. 20). The ratio of individuals in this experiment with 20:6 is already so intense that maximum reduction was achieved. As the period progressed, the difference between the control and the variants with the predator became greater. This is evidently a result of very effective suppression of the growth in the population of P. truncatus, by the predator.
T. nigrescens was able to develop well in both variants despite the very close ratios in individuals (Tab. 20). in the variant 20:10, reproduction of the predator was even slightly better. During the course of the experiment in both variants, there was a further narrowing down of the prey-predator ratio to a value of 1.3:1 although the initial value deviates considerably from natural conditions which are in the region of 7.5:1 (B�YE, 1988).
Tab. 20: Development of the population of Prostephanus truncatus, (P.t.) and Teretriosoma nigrescens (T.n.) on maize grains in relation to the initial ratio of individuals
| P.t.:T.n. | adult
P. truncatus after...days |
adult
T nigrescens after...days |
||||
| 50 | 70 | 110 | 50 | 70 | 110 | |
| Control | 290 a | 710 a | 888 a | -- | -- | -- |
| 20:10 | 26 b | 37 b | 48 b | 12 | 23 | 29 |
| 20:6 | 22 b | 38 b | 35 b | 8 | 16 | 26 |
Corresponding to how the population of the pest developed, the loss parameters in the presence of the predator were clearly reduced (cf. Tab. 21). The damage in the control after 50 days already amounted to 55.7% and, at this figure, was more than twice as high as in the other variants. At later dates, the damage in the presence of the predator was also significantly reduced.
With the progression of the experiment, loss in the control showed rapid increase and reached a figure of 63.3% after 110 days. In the variants with T. nigrescens, the increase developed in the same way, yet at a far lower level. after 110 days, the loss in this variant at 15% and 17%, was 76% and 73% lower than in the control.
Tab. 21: Damage and loss caused by Prostephanus truncatus, (P.t.) to maize grains in relation to the initial ratio of individuals in the presence of Teretriosoma nigrescens (T.n.)
| P.t.:T.n. | Damage
(%) after...days |
Loss (%) after...days |
||||
| 50 | 70 | 110 | 50 | 70 | 110 | |
| Control | 55,7 a | 83,9 a | 88,5 a | 14,7 a | 44,8 a | 63,3 a |
| 20:10 | 27,1 b | 47,4 b | 50,7 b | 4,9 b | 11,1 b | 17,2 b |
| 20:6 | 22,1 b | 48,8 b | 52,1 b | 4,4 b | 10,2 b | 15,2 b |
In this experiment, T. nigrescens proved to be a very effective antagonist of P. truncatus. In comparison to the results of experiments in Costa Rica, no considerable difference in the efficacy could be determined. With a prey-predator ratio of 2:1, the predator was able to reduce the population of the Larger Grain Borer in Togo to a greater extent than in Costa Rica; with a ratio of 3.3:1, it was vice versa. The striking fact observed, was the ability of P. truncatus, to develop better in the controls of the experiment in Togo than in Costa Rica, which could indicate favourable environmental conditions and or better fitness of the beetles (cf. NISSEN, 1989).