T. nigrescens was able to reproduce on some of the host species provided. As reproduction of the predator without hosts on plant substrate can be excluded (5.4), individual hosts must have been eaten for the development of the T. nigrescens progeny. Normally, in the experiments with a predator fewer host insects were found than in the controls without T. nigrescens. Statistically however, the growth of the host populations could only be suppressed by T. nigrescens in few monocultures. In experiments with the laboratory strain of L. oryzae and R. dominica on maize, the tests with predators even showed significantly more hosts than in the controls.
Presumably, many predator individuals of the F1 generation would have to develop to be able to have a significantly restricting effect on the host population. According to REES (1985) for example, one T. nigrescens larva eats approximately 60 P. truncatus larvae during its development into an imago. Some of the following examples will however show that this explanation alone is not sufficient.
In the R. dominica cultures on maize (Tab. 3.1 and 3.2), more progeny of T. nigrescens were found than in the P. truncatus cultures on cassava, sorghum and wheat (Tab. 1). Despite this, no growth-restricting effect could be proven on the R. dominica populations, whilst the P. truncatus populations on the substrates mentioned were significantly influenced by T. nigrescens.
Also, only few F1 individuals of the predator were counted in the O. surinamensis cultures on oats, cassava and maize. Despite this, a significant effect of T. nigrescens could be determined.
A strong influence of T. nigrescens on the host population can be explained using calculation examples, although only a few or no progeny of the predator were found. According to LELIVELDT (1990), one T. nigrescens imago eats 5.7 eggs or 4.9 larvae of P. truncatus per day on average. REES (1985) determined a value of 1.7 P. truncatus larvae. Since all 10 T. nigrescens imagines used were normally alive after 2 months (60 days), the imagines alone could eat 10 x 5.7 x 60 (3420) eggs or 10 x 4.9 x 60 (2940) P. truncatus larvae according to LELIVELDT (1990). According to Rees (1985), a value of 10 x 1.7 x 60 (1020) P. truncatus larvae would be produced for the duration of the experiment.
In this way, the T. nigrescens imagines could make a substantial contribution to the decimation of the host numbers because of their long life, although, according to REES (1985), they only eat half as many prey as the larvae of the predator.
Two concrete examples of these calculations are mentioned here. In the experiments with O. mercator (Tab. 3.7) and T. granarium on bran (Tab. 3.9) T. nigrescens did not reproduce. Despite this, significantly fewer hosts could be found in the tests with the predator than in the controls. Thus, the T. nigrescens imagines put in at the beginning of the experiment must have eaten the O. mercator and T. granarium individuals.
In contrast to the larvae, the adult T. nigrescens are not dependent on animal substances for nutrition but can also feed on plant substrates containing starch. If the predator imagines in the host cultures prefer plant nutrition rather than the prey offered, the growth of the host populations is influenced less than if the T. nigrescens of the P and F1 generations both hunt.
In this way, no influence of the predator on the host populations can be explained although, in comparison, a lot of T. nigrescens progeny were found in the tests. Examples of this are the two series of experiments with R. dominica on maize (Tab. 3.1 and 3.2).
It is less probable that the R. dominica population was able to compensate for the losses caused by T. nigrescens by increasing the rate of egg-laying.
The biology as well as the morphology and behaviour of the prey organisms certainly play an important role in answering the question as to whether T. nigrescens can make use of the species of beetle provided as a host.
The larvae of some Coleoptera species are very mobile. They are easily able to escape from the predator attacking them. The larvae of the tenebrionid beetle and some species of saw-toothed beetle were able to remain untouched by T. nigrescens in this way. Relatively immobile larvae on the other hand, like those of P. truncatus and other wood-borers and of T. stercorea, are easy victims for T. nigrescens.
Some species of larvae are well protected from enemies by their skeletonized cuticula or by hair and bristles. In contrast, the larvae of P. truncatus D. porcellus, R. dominica or T. stercorea are relatively soft and without hair, making them easier to injure than e.g. the larvae of A. diaperinus, the two species of Tribolium or the larvae of Dermestidae with dense hair.
The larvae of A. diaperinus, in addition, grow to a size which eliminates them as prey for T. nigrescens. The larvae of smaller species of beetle, e.g. those of O. surinamensis and O. mercator are, in comparison, still acceptable prey for T. nigrescens when they are fully grown and ready for pupating.
Some of the larvae and imagines offered as hosts are able to actively defend themselves against the threat of an enemy or even attack him. Most of the larvae of post-harvest pest beetles have strong mandibles presumably not only for grinding food but also for defending themselves against predators.
Like many other species of larder beetle (Dermestidae), the larvae of the Khapra beetle (T. granarium) has spiny hairs which it can use as an effective weapon when under threat (MA et al., 1978). Many species of tenebrionid beetles have a strong odour which could be a warning, or defensive secretion (allomone), and could be perceived by T. nigrescens in a similar way. The prey then is not so attractive to the predator and thus remains unharmed.
Apart from this, bad-tasting prey could deter the enemy from eating other specimens. It is known that bugs make use of this phenomenon to protect themselves from predators like birds, reptiles and amphibians.
The breeding biology of the host also has an effect on the ability of T. nigrescens to reproduce on the most varied species.
The Sitophilus species normally lay their eggs in a long hole they have bored themselves inside substrate grains. The entrance to the breeding hole is then sealed using a drop of transparent secretion (HEINZE, 1983). The larvae of the pest thus develop in concealment in a grain of wheat or maize. Because T. nigrescens has difficulty in biting substrate grains (5.3), the beetles can develop inside the hard shell of the grain undisturbed by predatory enemies.
This is presumably how the larvae of bruchid beetles remain safe from T. nigrescens.
In contrast to this, P. truncatus lays its eggs in groups in one of its boreholes inside a substrate grain without closing the entrance to the egg deposit. The diameter of the boreholes is large enough for T. nigrescens to be able to reach the eggs and later the larvae without difficulty whilst searching for prey.
The boreholes of the two other species of Bostrichidae (D. porcellus and R dominica) are smaller and access is difficult or impossible for T. nigrescens imagines.
If there is an insufficient number of suitable prey organisms, the female T. nigrescens presumably lay only few eggs. Thus, only few larvae of the predator are to be found in the populations which could enter the boreholes due to the diameter of their body and their longish shape, keeping the egg deposits of smaller corn borers relatively safe from the predator.
Apart from larvae which were ready to pupate (L2) and F1 imagines of T. nigrescens, there were only isolated eggs, egg-larvae or pupae of T. nigrescens to be found. The following behavioural patterns of the hosts and the predator could provide an explanation for these findings.
A number of the host species provided are latent predators. T. castaneum, for example, not only eats individuals of its own species but also the eggs and pupae of other species when food is scarce (HEINZE, 1983) or when over-population occurs. The eggs and pupae of T. nigrescens which existed could thus have been eaten by the hosts after the F1 generation of the hosts had hatched.
Another possibility is that the larvae from the T. nigrescens eggs laid at the beginning of the experiment hatched and developed into L2 undisturbed. This caused the pressure of competition for food to increase in the relatively small test jars to such an extent that cannibalism occurred. Eggs laid at a later time, egg-larvae and defenceless pupae could thus have become victims of their own species.
The T. nigrescens larvae which were already mature were thus unable to find suitable places to pupate in the over-populated jars. This could explain why mainly L2 of the predator were found in many experiments.
In the experiments with primary pests on maize, the tests with P. truncatus were the only ones showing more F1 imagines of T. nigrescens than L2.
According to LELIVELDT (1990) and REES (1985), the T. nigrescens larvae crawl into hollowed-out maize grains to pupate. P. truncatus seems to be the only host which was able to hollow out the maize grains in such a way that the larvae found an ideal refuge where they could pupate. The feeding and breeding holes of R. dominica, D. porcellus and the Sitophilus members were evidently too small or were unsuitable for other reasons for providing the mature T. nigrescens larvae with a refuge. The assumption that sorghum and wheat grains could restrict or prevent the predator's larvae from pupating due to their small dimensions was supported by the results of several series of experiments.
In P. truncatus populations on sorghum and wheat as well as in S. oryzae populations on wheat, chiefly L2 of the predator could be observed. Even in O. surinamensis populations on oat flakes the larvae of the predator evidently found it difficult to find a place to pupate.
The experiments with secondary pests contained coarse meal as well as maize. Coarse maize meal can be used by the T. nigrescens larvae to build pupating cases. This could account for the comparatively high number of F1 imagines of the predator in the tests with O. mercator, O. surinamensis and T. stercorea.
The findings presented here clearly show that T. nigrescens tended to accept the species of Coleoptera offered more as prey organisms in the presence of P. truncatus than in the monocultures. Similarly, many host populations in the monocultures on maize remained uninfluenced by T. nigrescens whilst the number of specimens in the mixed cultures were significantly reduced by the predator (Fig. 14). REES (1987) and LELIVELDT (1990) were also able to observe this phenomenon.
Fig. 14: Influence of T. nigrescens on the development of pest populations on maize
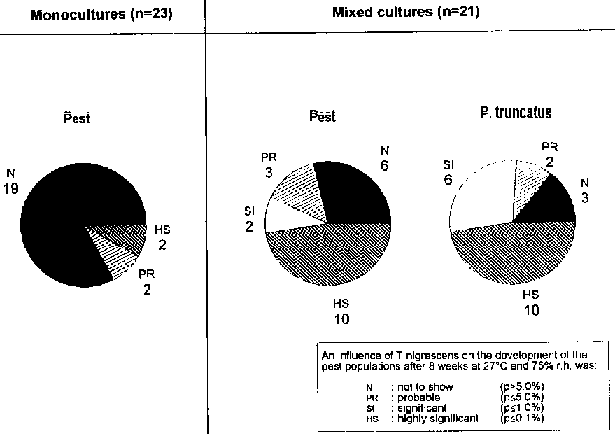{kind=link}
The following correlations could, according to B�YE (1988), have provided the cause for these findings. T. nigrescens can sense its prey over a long distance and actively search for it (REES et al, 1990; REES, 1990) using the aggregation pheromone of P. truncatus. This pheromone of the Bostrichida presumably works on T. nigrescens with a kairomone effect and stimulates the hunting and eating behaviour of the predator in the mixed cultures. Expecting to encounter its favourite prey P. truncatus whilst eating, the eggs and larvae of other host species in the test jar are also sought out and eaten.
In most cases, however, it remains uncertain whether T. nigrescens made use of the individuals of the Coleoptera species in the test jars as prey or just killed them. The findings from two series of experiments, however, indicate that T. nigrescens ate the hosts.
In the mixed cultures with T. castaneum and T. confusum from Togo (Tab. 3.5 and 3.6), no influence on the P. truncatus population could be determined while the number of Tribolium imagines was significantly reduced by T. nigrescens. In the former case, P. truncatus was not able to reproduce, and in the second case only insignificantly. An average of nine to ten T. nigrescens offspring could be found in these experiments. Since in these cases there were no or very few P. truncatus individuals as prey, it can be assumed that the predator made use of the second host species as a source of nourishment and reproduced on this.
T. nigrescens multiplied better in many mixed cultures than in pure P. truncatus populations of a comparable size. REES (1987) and LELIVELDT (1990) were also able to observe a high rate of reproduction for the predator in these kinds of mixed cultures.
The host insects could have laid more eggs under the pressure of competition to preserve their own species. T. nigrescens would thus have had more suitable prey organisms than, for example, in the monocultures with P. truncatus. Perhaps eating several species of prey also contributes to better nourishment of the predator which is then reflected in an increased rate of reproduction.
According to LELIVELDT (1990), T. nigrescens also attacks freshly hatched P. truncatus imagines when food is scarce. In two cases it could be proven that T. nigrescens also eats imagines belonging to other species of beetle.
R. dominica was not able to use cassava as a breeding substrate. In comparison to the controls, the experiments with T. nigrescens contained a highly significant lower number of adult hosts (Tab. 3.1). This allows the conclusion that the predator also accepted adult R. dominica imagines as prey organisms. Whether T. nigrescens killed the adult R. dominica or only ate the beetles after death, cannot be judged as all the insects were frozen prior to evaluation. Thus it is not known how many of the R. dominica imagines in the controls were dead at the end of the experiment.
Cannibalism among R. dominica imagines, induced by the presence of T. nigrescens, is unlikely to be another explanation.
A comparable case occurred in the mixed cultures with P. ratzeburgi (Tab. 3.5). The P. ratzeburgi imagines did not reproduce in the controls. In the experiments with the predator, a highly significant lower number of adult hosts of the P generation were found. T. nigrescens consequently accepted adult P. ratzeburgi as prey organisms in the mixed population of P. ratzeburgi and P. truncatus
In six other series of experiments (with D. porcellus and O. surinamensis on cassava, and in the mixed cultures with D. porcellus, O. mercator, C. dimidiatus and T. granarium on maize) the number of host imagines counted was far lower in the experiments with T. nigrescens than the number originally inserted. In each of these cases the reproductive activities observed for the hosts were mostly only low.
The conclusion that T. nigrescens had accepted the imagines of all of the host species as prey in the mixed cultures is probable but cannot be proven. It is not known how many of the adult hosts found in the experiments without T. nigrescens belong to the parent generation. It could be true that only few of the originally inserted host imagines remained healthy also in the controls. A comparison of results in the experiments with predators with the control results is thus not possible.
Table 4: The ability of T. nigrescens to reproduce on various stored product pest Coleoptera and the influence of the predator on the development of these populations
The average number of T.nigrescens progeny was between:
| *: | 0.2 and 1.0 | #: 5.4 and10.0 |
| **: | 1.2 and 3.0 | ## :10.2 and20.0 |
| ***: | 3.2 and 5.8 | ###: 20.2 and36.8 |
| -: no progeny of T.nigrescens | •: not done | |
| /: result first experiment / second experiment | ||
An influence of T.nigrescens on the development of the pest populations was:
N : not to show (p > 5.0%)
PR : probable (p<5.0%)
SI : significant (p<1.0%)
HS : highly significant (p<0 1%)
{} : result for P.truncatus populations
| MONOCULTURES | MIXED CULTURES On maize with {P.trun.} |
||||||||
| Pest | Breeding Substrate |
Maize |
Cassava |
||||||
| D.porc.T | * | N | ** | N | ** | HS | # | HS | {HS} |
| R.domi. | -/** | N/N | ***/*** | N/N | * | HS | ### | PR | {HS} |
| S.gr.af. | * | N | * | N | • | • | ### | N | {HS} |
| S.gr.gr. | * | N | * | N | • | • | ## | SI | {SI} |
| S.oryz. | *** | N | ** | N | • | • | ### | HS | {HS} |
| S. zeam. | • | • | * | N | * | N | ### | PR | {PR} |
| A.diap.T | • | • | * | N | • | • | • | ## | {HS} |
| L. oryz. | - | N | - | N | • | • | ### | N | {HS} |
| L.oryz.T | * | SI | - | N | • | • | ### | HS | {HS} |
| P.ratz. | - | N | * | N | • | • | ## | HS | {SI} |
| P. subd | . - | N | • | • | • | • | • | • | |
| T.cast. | - | N | - | N | - | N | # | HS | {N} |
| T.conf. | - | N | * | N | - | N | ## | HS | {SI} |
| T.conf.T | - | N | */- | PR/N | - | N | # | ST | {N} |
| A.adve.T | - | N | - | PR | • | • | ## | N | {HS} |
| C.pusi. | - | N | - | N | • | • | ### | HS | {SI} |
| O.merc. | - | HS | ** | HS | • | • | ## | HS | {HS} |
| O.suri. | *** | SI | ** | HS | * | SI | ## | N | {PR} |
| C.dimi. | - | PR | - | N | • | • | # | HS | {ST} |
| C.dimi.T | - | PR | - | N | • | • | # | N | {N} |
| T .gran. | - | SI | - | N | • | • | ### | PR | {HS} |
| T.ster. | ** | N | ** | N | ** | SI | # | N | {SI} |
Evaluation after 8 weeks at 27�C and 75% r.h. (mw, n=5); key to the abbreviations used for the pests Tab.2
Apart from the host insects themselves, the nutritive medium for the hosts also seems to play an important role in the development of T. nigrescens. Reproduction of T. nigrescens could be observed (Tab. 4) in 61% of all series of experiments on maize (n=23; without the series containing P. truncatus In contrast, only in 38% of the experiments (n=21; without the series containing P. truncatus on the usual nutritive substrates like bran, wheat, oat flakes and sorghum could offspring of T. nigrescens be found.
T. nigrescens only reproduced on the O. mercator cultures when the insects were kept on maize and did not reproduce on bran.
Only in 4 of the total 10 series of experiments with R. dominica on sorghum could progeny of T. nigrescens be found. T. nigrescens, in contrast, reproduced successfully on maize in all 10 tests with R. dominica as the host species.
In comparison to maize, the R. dominica and O. mercator imagines were able to reproduce far better on the substrate they were accustomed to. In the first series of experiments with R. dominica on sorghum for example, 7.5 times more adult R. dominica were counted than in the tests started at the same time on maize. Consequently, there were considerably more specimens of prey available to the predator in the tests on sorghum. Despite this, offspring of T. nigrescens could only be found on maize during this series of experiments.
From this, it can be concluded that the ability of T. nigrescens to reproduce can depend not only on the number of prey organisms available, but the nutritive medium maize also has a key function in the development of T. nigrescens.
R. dominica, S. zeamais, T. castaneum and both strains of T. confusum were unable to reproduce on cassava. P. truncatus D. porcellus and O. surinamensis showed only a low rate of reproduction on the dried tubers.
The assumption that residues of insecticides could have been attached to the cassava roots from Togo and could have had a restricting effect on the beetles' ability to reproduce, became improbable according to what HELBIG (1992) said. His information showed that dried cassava roots are normally not treated with insecticides.
In addition, T. stercorea was able to reproduce better on the cassava roots from the same lot than on any other substrate. Many of the imagines of the other species tested and originally inserted, were still alive on the cassava 2 months after the experiments began.
Most of the species of beetle used for the investigations on cassava came from laboratory cultures which had been bred on other nutritive media for many years. It is, however, improbable that the populations have lost their original ability to reproduce on cassava due to this fact.
Cassava thus seems to be a good nutritive substrate for most species of pest beetle, yet unsuitable as a breeding substrate for the Coleoptera species.
In many cases P. truncatus was only able to multiply weakly when another host species was present.
It was evident that the Sitophilus species dominated over the Bostrichida in the experiments with S. zeamais and S. oryzae. The weevils/snout beetles, which like P. truncatus are primary pests, successfully competed with P. truncatus for food and breeding spaces.
That the Sitophilus species are not superior in general, is shown by the series of experiments with 5. granarius granarius The ratio of individuals in both populations was relatively balanced. In the mixed cultures with 5. granarius africanus, P. truncatus was dominant. P. truncatus was able to assert itself against the two other Bostrichidae, D. porcellus and R. dominica in the mixed cultures.
In the mixed cultures with P. truncatus and one of the tenebrionid or Cucujidae and in one of the two strains of C. dimidiatus and T. stercorea, both host populations extensively developed badly in comparison to the findings in monocultures on maize.
Two experiments carried out by B�YE (1988) show that the results of the investigations on loose maize grains cannot be simply transferred to the situation with stored maize cobs. He, too, was able to prove that loose maize grains provided S. zeamais with an advantage over the P. truncatus populations. If, on the other hand, the two species of beetle were put onto maize cobs, this boosted the development of the P. truncatus populations.
In the monocultures as well as in the mixed ones with secondary pests, coarse maize meal was added to the maize grains as a substrate. Improved reproduction of the secondary pests in the presence of P. truncatus as a primary pest could thus neither be expected nor observed.
T. nigrescens seems to have accepted the wild strain of L. oryzae from Togo as a prey more than the laboratory strain.
Whilst the laboratory strain was not influenced by T. nigrescens in any of the experiments, the number of L. oryzae imagines belonging to the wild strain from Togo was significantly reduced by the predator on bran as well as in the mixed culture.
Vital differences between the strains of T. confusum and C. dimidiatus bred in the laboratory for many years and the wild strains from Togo as hosts for T. nigrescens could not be determined.
A suitable method of investigating the host specificity of T. nigrescens was sought in preliminary experiments (P�SCHKO, 1992 b). The results varied depending on the procedure. In experiments with O. surinamensis, neither an ability of T. nigrescens to reproduce nor a restricting effect on the growth of the populations could be observed, in contrast to later findings.
For this reason, a uniform method was applied in these investigations allowing a direct comparison between the suitability of the species of pest beetle as hosts for T. nigrescens.
Contradictory results - also within these uniform experiments - clearly show that many parallel experiments and repetitions of complete series of experiments are necessary before precise statements on the host specificity of T. nigrescens can be made. T. nigrescens, for example, reproduced in a series of experiments with R. dominica cultures on sorghum, yet in the repeat experiments no offspring of T. nigrescens could be found in the tests (Tab. 3.1).
With other experimental methods, isolated F1 individuals of T. nigrescens could certainly be found also on strains of beetles not appearing here as hosts.
When additionally taking the long life expectancy of the T. nigrescens imagines into consideration and the ability of the females to lay eggs over several months (3.2), a reproduction of T. nigrescens in the laboratory would also be presumably be possible on host species other than P. truncatus.
All findings on the host specificity of T. nigrescens presented here are based on laboratory experiments. It can be assumed that the results can only be transferred to natural situations with some limitations.
In this respect for example, the insects were kept under constant climatic conditions favourable to their development. The species of pest insects in a store may be relatively well protected from seasonal climatic fluctuations due to the micro-climate inside the maize cobs stored with husks, but despite this, extremely low or too high temperatures, or too dry or damp conditions could occur which could have various effects on the development of the species of beetle inside the maize cobs.
Neither the predators nor the hosts were able to escape from the system set up, for example, either to go in search of more preferred sources of nutrition or to escape over-population. In a traditionally constructed maize store in contrast, it is possible at any time for the insects to move off or for other species of pests or useful insects to immigrate.
Laboratory experiments will, however, continue to be indispensable in being able to understand and study the complex relationship between the predator T. nigrescens and its host P. truncatus step by step.
The results of these experiments combined with the findings of 3.3 highlight the dominant role of P. truncatus as a host for T. nigrescens.
Multiplication of T. nigrescens only took place in P. truncatus cultures on maize during the period of experimenting.
Reproduction activity of T. nigrescens could be observed on all the substrates on which P. truncatus reproduced.
T. nigrescens was also able to reproduce in the P. truncatus cultures when the medium used as a breeding substrate for P. truncatus was less suitable and when only a few hosts were available, as was the case, for example, on cassava and wheat.
T. nigrescens significantly restricted the growth of the P. truncatus populations on all breeding substrates determined for the pest.
In the mixed cultures where one each of the species of beetle selected was kept with P. truncatus, T. nigrescens normally showed a high rate of reproduction. In addition to suppressing the P. truncatus populations, T. nigrescens also suppressed the growth of other populations of pest species.
The data presented here provides a basis suggesting that after release, T. nigrescens will successfully reproduce and spread in Togo and West Africa. In natural conditions, T. nigrescens could only be observed in connection with P. truncatus B�YE 1988; REES, 1990). Consequently, T. nigrescens will be found exclusively in maize stores infested by this pest.
Infestation by primary pests attracts other species of pest, such as tenebrionid and saw-toothed beetles which are not able to damage intact maize grains by themselves. It can be assumed that T. nigrescens, as in the laboratory experiments, primarily accepts P. truncatus individuals as prey. However, apart from this, eggs and larvae of other species of pest are eaten in the presence of P. truncatus. T. nigrescens will be able to reproduce well in this natural constellation of species consisting of several pest-beetle populations. If the density of the predator in the store becomes too great, giving rise to competition for food among the predator populations, or the supply of nourishment for the predator becomes worse, the adult T. nigrescens will emigrate in order to search for another maize store infested by the host it prefers, P. truncatus, with the aid of the kairomone effect.
In this way, it can be assumed that biological control of P. truncatus using T. nigrescens will be successful and will result in a reduction in damage and losses to stored maize in Africa.
4.2 Host specificity of Teretriosoma nigrescens among species of Lepidoptera storage pests
4.2.1
Introduction
4.2.2
Method
4.2.3 Results and Conclusion
Besides approximately 60 species of Coleoptera, there are in the region of a dozen species of Lepidoptera which play an economically important role in regard to insect storage pests (WOHLGEMUTH, 1991).
Investigations on the host specificity of T. nigrescens were carried out with the 4 most significant species of pyralid moths, Ephestia cautella, E. elutella, E. kuehniella and Plodia interpunctella, the rice moth Corcyra cephalonica and the angumois grain moth Sitotroga cerealella (Tab. 2).
First of all, moth eggs were offered to T. nigrescens as prey in small Petri dishes without any substrate.
In the second phase of the investigations, the moth eggs were put onto a nutritive medium in a breeding jar. The influence of T. nigrescens on the development of the moth populations could be examined in this way.
Finally, the development of a mixed population from eggs of varying species of moth was observed in the presence of P. truncatus with and without the influence of T. nigrescens.
The T. nigrescens imagines ate the moth eggs offered to them without a substrate.
The predator had no restricting effect on the development of the moth populations on substrate.
There was also no evidence of a reducing effect of T. nigrescens on the number of moths developing in the presence of its host P. truncatus
The results clearly show that T. nigrescens can make use of the eggs of moth storage pests for nourishment; a mechanism allowing targeted search for these prey organisms, however, does not seem to exist.
Due to these findings, further investigations on the ability of T. nigrescens to reproduce and multiply with Lepidoptera as hosts were not carried out.