4 Interactions entre Prostephanus truncatus et Teretriosoma nigrescens
Table
des mati�res - ![]() Pr�c�dente
- Suivante
Pr�c�dente
- Suivante![]()
4.1 Influence de teretriosoma nigrescens dans
des conditions proches de la r�alit�
4.2 Comparaison de l'efficacit� de Teretriosoma
nigrescens au Togo et au Costa Rica
4.1 Influence de teretriosoma nigrescens dans des conditions proches de la r�alit�
4.1.1 Mat�riels et m�thodes
4.1.2 Populations de ravageurs et d'insectes
utiles
4.1.3 Etat des dommages et pertes
Avant de l�cher le pr�dateur au Togo, il s'agissait en l'occurrence d'�tudier dans des conditions de quarantaine l'influence de l'insecte utile import� T. nigrescens sur P. truncatus, le ravageur introduit dans le pays. A cette fin, on a install� des greniers � mais du type de ceux utilis�s habituellement dans le sud du Togo, en les isolant dans une cage de toile m�tallique destin�e � emp�cher toute immigration ou �migration des insectes. De mani�re � rassembler tout de m�me dans ces greniers un �ventail de ravageurs analogue � celui qui appara�t dans le contexte naturel, le mais, avant fermeture de la cage et introduction du pr�dateur, a �t� expos� durant une p�riode de 4 mois � l'infestation naturelle par les insectes nuisibles.
En ao�t 1989, 6 greniers exp�rimentaux du type traditionnel "Ebli-va" ont �t� �rig�s sur une plate-forme de b�ton (6 m sur 10) pr�par�e � cette intention (fig. 17). Sur le b�ti inf�rieur en bois des greniers (80 cm de haut), lequel comportait quatre pieds, on a install� une plate-forme (120 cm de long, 120 cm de large) construite dans les esp�ces de bois couramment utilis�es dans la r�gion. L'emmagasinage du mais (environ 200 kg d'une esp�ce locale � grains jaunes par grenier) a eu lieu les 23 et 24 ao�t 1989. Les 15 et 16 novembre 1989, les �pis de mais ont �t� retir�s de la totalit� des greniers et syst�matiquement m�lang�s. Les greniers ont �t� ensuite reconstitu�s. Cette op�ration avait pour but de garantir une r�partition homog�ne des insectes ravageurs dans tous les greniers.
Cage exp�rimentale
Cette cage en toile m�tallique (5 m de largeur, 9 m de longueur et 2,50 m de hauteur) (fig. 17), �tait partag�e en deux moiti�s dans le sens longitudinal, chaque demi-espace renfermant quatre cellules dispos�es les unes derri�re les autres. Il s'agissait dans chaque cas de trois cellules exp�rimentales (2,50 m sur 2,50 m) et d'une entr�e (1,50 m sur 2,50 m) servant de sas. Les deux demi-espaces n'�tait pas reli�s entre eux. A l'int�rieur de chaque demi-espace, chaque cellule �tait reli�e � la suivante par une porte. Chaque moiti� de cage a �t� utilis�e pour les trois r�p�titions effectu�es sur une m�me variante. Le b�ti de la cage �tait en fer plat, sur lequel on avait appliqu� un produit antirouille. La toile m�tallique en acier fin (mailles de 0,5 mm de large) �tait tendue entre les fers plats. Pour en garantir l'�tanch�it� absolue, cette toile avait �t� tremp�e dans un agent de densification des plastiques. L'�coulement d'eau �tait assur� s�par�ment pour chaque moiti� de la cage par des siphons aboutissant � des conteneurs de r�cup�ration ferm�s en b�ton.
Introduction de T. nigrescens
Le transfert des T. nigrescens de l'Institut f�d�ral de biologie (BBA) de Berlin � Lom� s'est d�roul� le 17 novembre 1989 dans le respect des directives concernant la quarantaine. Pr�alablement � ce transfert, 5 � 6 g�n�rations du pr�dateur lequel avait �t� collect� au Costa Rica en novembre 1988 - avaient �t� mises en observation en quarantaine au BBA, multipli�s et soumis � un examen de d�pistage d'agents pathog�nes. Les animaux ne pr�sentaient aucun agent pathog�ne ni antagoniste. L'introduction du pr�dateur dans les cages, � raison de 2 000 adultes par r�p�tition, a eu lieu le 18 d�cembre 1989.
Echantillonnage et �valuation
Les premiers pr�l�vements d'�chantillons de ma�s ont �t� effectu�s en ao�t 1989, aussit�t apr�s l'emmagasinage. Apr�s deux autres �chantillonnages de 100 �pis � un mois d'intervalle, on a observ� une pause, pendant laquelle on a m�lang� les �pis, �rig� de nouveaux greniers et construit la cage au-dessus des greniers. Apr�s la fermeture de cette cage, mais avant l'introduction de T. nigrescens, on a recommenc� � pr�lever r�guli�rement des �chantillons. Ces �chantillons ont �t� plac�s dans une caisse m�tallique et fumig�s � l'hydrog�ne phosphor� dans l'entr�e de la cage durant au moins une semaine avant d'�tre soumis � �valuation. L'�valuation a �t� effectu�e individuellement pour chacun des 100 �pis de l'�chantillon. Pour ce qui est du calcul des pertes, on s'est servi de la m�thode du comptage et du pesage (chapitre 2.1.2).
Nous nous sommes limit�s dans la pr�sentation des r�sultats aux principaux ravageurs primaires et param�tres de pertes. On trouvera les r�sultats complets chez HELBIG (1993).
4.1.2 Populations de ravageurs et d'insectes utiles
L'�tude des interactions entre P. truncatus et T. nigrescens, de m�me que l'�volution d�mographique qui en d�coule pour le ravageur constituent l'objet essentiel des recherches pr�sent�es ici. Au-del�, il �tait toutefois int�ressant d'observer l'�volution d�mographique des autres ravageurs au cas o� il existerait �galement entre eux et le pr�dateur des rapports particuliers.
P. truncatus
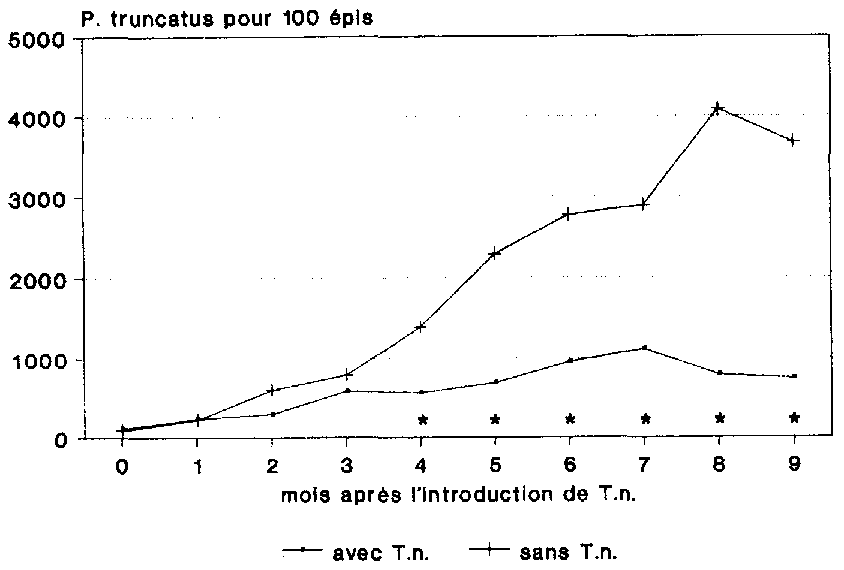
L'influence du pr�dateur sur l'�volution d�mographique du ravageur s'est manifest�e au bout de 2 mois d�j� apr�s son introduction, et cela � travers le nombre de P. truncatus adultes trouv�s (fig. 18). A cette date pr�coce, il n'y avait qu'une diff�rence relativement minime et pas encore significative de 301 col�opt�res entre les deux variantes. Au cours des mois suivants, on a enregistr� dans la variante sans T. nigrescens une forte croissance de la population de P. truncatus, qui s'est traduite au bout de 8 mois par un maximum de 4 086 adultes pour 100 �pis.
Compar�e � l'�volution d�mographique observ�e dans le t�moin, le ravageur a connu dans la variante impliquant T. nigrescens une croissance d�mographique fortement limit�e. Le nombre maximum de P. truncatus adultes, I 1 17 col�opt�res, a �t� obtenu au bout de 7 mois. A l'issue des 9 mois d'essai, le nombre de P. truncatus adultes �tait, sous l'influence du pr�dateur, inf�rieur de 79,8 % (760) � celui trouv� dans le t�moin (3 670). La mise en oeuvre de T. nigrescens a donc permis de circonscrire tr�s efficacement la croissance d�mographique du grand capucin du mais. Du fait que seuls les animaux adultes ont �t� recens�s dans le cadre de cet essai, l'efficacit� du pr�dateur ne s'est toutefois manifest�e nettement qu'avec un certain d�calage temporel.
T. nigrescens
A la suite de son transfert du Costa Rica au Togo, via Berlin, le pr�dateur a �t� introduit dans un nouvel habitat. Il r�gne dans ce nouvel habitat des conditions environnementales diff�rentes de celles de sa r�gion d'origine, l'Am�rique centrale. L'essai d�crit s'�tant d�roul� dans une cage de toile m�tallique ferm�e, le nombre des facteurs environnementaux �tait n�cessairement limit�. On peut par cons�quent consid�rer ici les conditions climatiques comme facteur d'influence essentiel.
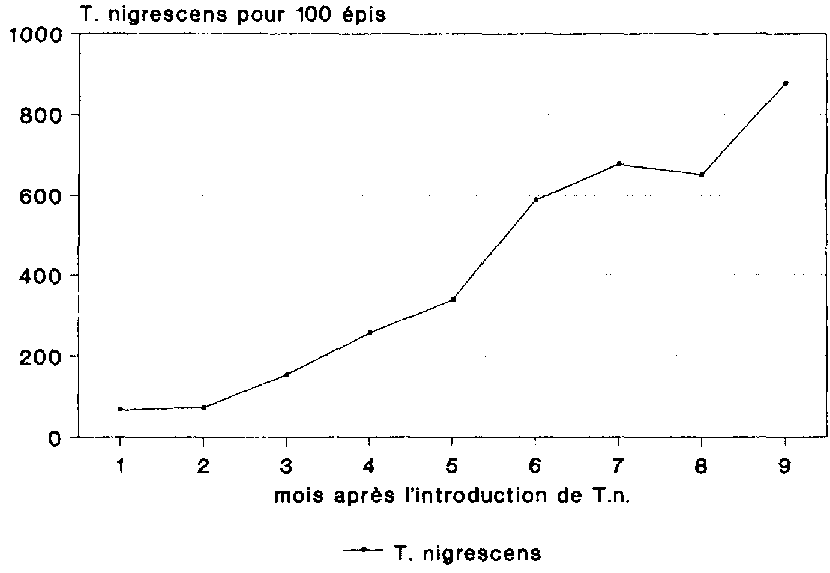
T. nigrescens n'a eu visiblement aucune difficult� majeure � s'adapter � son nouvel environnement (fig. 19). La population n'a cess� de s'accro�tre pendant toute la dur�e de l'essai, parvenant au bout de 9 mois � une densit� moyenne de 876 individus adultes pour 100 �pis.
Si l'on consid�re � pr�sent l'abondance de T. nigrescens en comparaison de la population de P. truncatus, on arrive au bout de 9 mois � un rapport proie-pr�dateur de 1 contre 1,2. Bien que l'on ait constat� d�s le d�but de cet essai un rapport proie-pr�dateur de 3,6 contre 1, donc tr�s �troit compar� aux recensements effectu�s au Costa Rica en milieu naturel (cf. B�YE, 1988), ce rapport s'est encore resserr� au cours du stockage.
S. zeamais
S. zeamais doit �tre consid�r� comme le principal ravageur � part P. truncatus, puisque aussi bien il est responsable pour une bonne part des dommages et pertes caus�s aux stocks de ma�s. Il est donc int�ressant, du point de vue de la physionomie des param�tres de pertes, d'observer son �volution d�mographique. Dans l'optique d'effets secondaires �ventuels li�s � la mise en oeuvre de T. nigrescens, il est par ailleurs important de comparer le d�veloppement de la population de S. zeamais avec et sans pr�dateur.
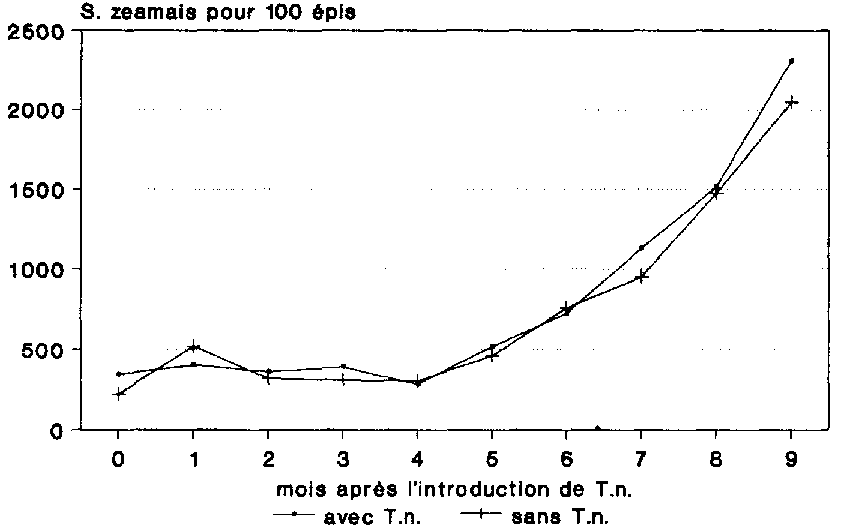
Jusqu'au quatri�me mois de stockage, la densit� de population de S. zeamais est demeur�e � son niveau initial de 300 � 400 col�opt�res environ pour 100 �pis (fig. 20). Ce n'est qu'au bout du cinqui�me mois de stockage que la population a commenc� d'augmenter de fa�on sensible. Pendant les mois qui ont suivi, les populations des deux variantes ont connu une progression presque parall�le, atteignant apr�s 9 mois d'essai une densit� respective de 2 048 et 2 308 col�opt�res pour 100 �pis. Cet essai n'a pas permis d'�tablir une influence quelconque de T. nicrescens sur l'�volution d�mographique de S. zeamais.
Ravageurs secondaires
Eu �gard � d'�ventuels effets secondaires li�s � la mise en oeuvre de T. nigrescens, il convient �galement de prendre en consid�ration les populations de ravageurs secondaires. L'�tude et l'interpr�tation de leur �volution d�mographique impliquent toutefois que l'on tienne compte de leur d�pendance vis-�-vis des ravageurs primaires. Dans le cas du stockage du mais sous forme d'�pis, notamment, l'infestation par les ravageurs primaires constitue une condition indispensable � une exploitation �ventuelle du milieu nutritif par les ravageurs secondaires.
P. truncatus produit une quantit� de farine de forage consid�rable, laquelle sert de nourriture de base aux ravageurs secondaires. Leur d�veloppement est par cons�quent subordonn� � la quantit� de farine de forage produite et d�pend indirectement de l'importance de la population du grand capucin du mais. L'influence de T. nigrescens sur la population de P. truncatus et, par suite, sur la quantit� de farine de forage, a par l� m�me un caract�re indirect. Vu les conditions de r�alisation de l'essai qui nous occupe, il n'�tait pas possible de distinguer ce type d'influence d'une �ventuelle influence directe.
Les greniers exp�rimentaux dispos�s dans la cage de toile m�tallique �tant expos�s � l'infestation naturelle, les ravageurs secondaires qui s'y sont manifest�s �taient les m�mes que dans les greniers des paysans, � savoir Carpophilus spp., C. quadricollis, T. castaneum, Cryptolestes spp., P. subdepressus et O. surinamensis. Concernant Carpophilus spp., C. qundricollis et O. surinamensis, on a constat� aussi bien dans les greniers avec et sans le pr�dateur une �volution d�mographique pratiquement identique. Pour ce qui est des Cryptolestes spp. et de T. castaneum, on a observ� un l�ger recul de la densit� dans les greniers o� T. nigrescens �tait pr�sent. P. subdepressus, en revanche, accusait une nette r�duction de population.
S'agissant des Cryptolestes spp, et de P. subdepressus, il est possible qu'il y ait eu une influence indirecte de la part de T. nigrescens. Dans la variante impliquant le pr�dateur, la population du grand capucin du ma�s avait nettement r�gress� (voir plus haut), entra�nant du m�me coup une diminution de la farine de forage produite. Les ravageurs secondaires rencontraient donc dans cette variante des conditions nutritives moins favorables. Cette hypoth�se pourrait �galement s'appliquer � T. castaneum. Ce ravageur �tant aussi connu comme pr�dateur, on peut supposer l� encore que la l�g�re r�duction de la densit� d�mographique observ�e dans la variante avec pr�dateur �tait due � une moindre abondance de proies.
4.1.3 Etat des dommages et pertes
En compl�ment de l'�tude de l'�volution d�mographique des ravageurs, on a �galement proc�d� � une analyse de l'�tat des dommages et des pertes, analyse destin�e � fournir certaines indications quant � l'efficacit� de T. nigrescens au niveau des pertes caus�es par P. truncatus. Les r�sultats n'en sont toutefois pas enti�rement fiables dans la mesure o� l'essai s'est d�roul� seulement dans des conditions proches de la r�alit� et non pas dans le contexte naturel.
Dommages
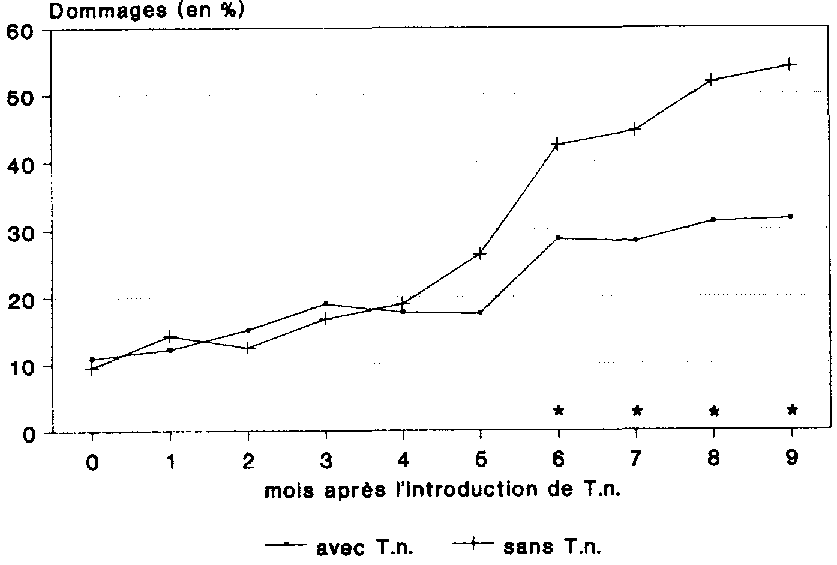
Au moment de l'introduction du pr�dateur T. nigrescens dans les greniers � ma�s, les dommages s'�tablissaient aux alentours de 10 % dans les deux variantes (fig. 21). Au cours des quatre mois suivants, on a enregistr� une l�g�re progression des dommages, qui atteignaient alors environ 20 % dans les deux variantes, une nette diff�rence apparaissant par la suite.
Dans la variante sans pr�dateur, on a enregistr� une forte augmentation des pertes qui s'est maintenue jusqu'� la fin de la p�riode d'observation. A l'issue de la p�riode de stockage de 9 mois, les dommages caus�s dans le t�moin s'�levaient � 54,1 %.
Dans les greniers � ma�s o� l'on avait mis T. nigrescens en oeuvre, on a �galement constat� une augmentation des dommages, beaucoup moins accentu�e cependant que dans le t�moin. De 20 % au cinqui�me mois de stockage, les dommages sont pass�s � 30 % environ au cours du sixi�me mois. Les dommages se sont maintenus � ce niveau pendant la p�riode qui suivi, pour s'�tablir � 31,6 % � la fin des 9 mois d'essai, ce qui, par rapport au t�moin, repr�sente une r�duction statistiquement significative de 41,6 %.
Pertes
Parties d'un chiffre de 3 %, les pertes n'ont progress� que faiblement dans les deux variantes au cours des trois premiers mois qui ont suivi l'introduction du pr�dateur, passant alors � 7 % (fig. 22). Par la suite, les pertes enregistr�es dans les greniers � ma�s o� le pr�dateur �tait absent ont augment� beaucoup plus fortement que dans la variante impliquant T. nigrescens. Cette �volution s'est poursuivie jusqu'� la fin de la p�riode d'observation. Au bout de 9 mois d'essai, on a relev� dans le t�moin des pertes de MS de 37,2 %.
Dans les greniers � mais o� l'on avait fait intervenir le pr�dateur, on a �galement constat� une augmentation de pertes, inf�rieure toutefois � celle de la variante de r�f�rence, et qui s'est amenuis�e au fur et � mesure que la p�riode d'essai s'avan�ait. Au bout de 9 mois d'essai, on a relev� des pertes de 19,9 % apr�s intervention de T. nigrescens, ce qui repr�sente 46,5 % de moins que dans les greniers � mais o� le pr�dateur �tait absent.
Influence de T. nigrescens sur la structure des dommages
On s'est livr� l� encore dans le cadre de cet essai � une analyse de la structure des dommages. Cette structure peut servir d'orientation pour d�terminer les pertes de mais consommable subies par les paysans � la suite d'une infestation d'insectes (voir chap. 2.6). La r�partition en cat�gories a �t� effectu�e en fonction du bar�me indiqu� au chapitre 2.1.2. Les �pis pr�sentant plus de 75 % de grains endommag�s ont �t� class�s dans la cat�gorie de dommages 5.
Dans les greniers � mais o� le pr�dateur �tait absent, la part d'�pis de la cat�gorie 5, qui �tait de 7 % en d�but d'essai, a connu une augmentation permanente pendant toute la dur�e de l'essai (tabl. 18). Au bout de 9 mois d'essai, 56 % des �pis �taient � ranger dans la cat�gorie 5. En partant de l'hypoth�se que les �pis de la cat�gorie 5 ne sont plus livr�s � la consommation, cela signifie une perte de ma�s consommable de 56 %.
Dans les greniers o� T. nigrescens avait �t� mis en oeuvre, le nombre d'�pis de la cat�gorie 5 a connu au cours des premiers mois de stockage une �volution parall�le � celle de la variante t�moin (tabl. 19). Il s'agit l� d'un ph�nom�ne d�j� observ� pour les autres param�tres de pertes. On a assist� � partir du cinqui�me mois � des diff�rences entre les deux variantes, le nombre des �pis de la cat�gorie 5 ayant augment� beaucoup plus lentement en pr�sence du pr�dateur au cours de la p�riode qui a suivi.
Il est frappant de constater la forte augmentation du pourcentage d'�pis de la cat�gorie 5 du cinqui�me au sixi�me mois suivant le d�but de l'essai. Ce bond en avant se refl�te �galement dans l'�volution des dommages, de m�me que dans celle des pertes et de la production de farine de forage (voir plus haut). S'il n'y a pas eu de d�veloppement parall�le des populations de P. truncatus, celle de S. zeamais a connu en revanche une forte croissance au cours de cette p�riode. Il est par cons�quent possible que ce soit ce ravageur qui est � l'origine des dommages (cf. chapitre 4.1.2). A l'issue de la p�riode d'essai de 9 mois, 31 % des �pis �taient � classer dans la cat�gorie 5 apr�s mise en oeuvre du pr�dateur. Les pertes de mais consommables �taient ainsi inf�rieures de 44 % � la variante sans pr�dateur.
Tabl. 18: Structure des dommages caus�s aux �pis sans intervention du pr�dateur Teretriosoma nigrescens (pourcentage d'�pis endommag�s sur un total de 300, � chaque date consid�r�e et dans les diverses cat�gories de dommages)
| Cat�gorie
de dommages |
Mois apr�s l'introduction du pr�dateur |
|||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | 61 | 61 | 60 | 54 | 53 | 44 | 36 | 34 | 25 | 27 |
| 2 | 27 | 23 | 25 | 24 | 24 | 24 | 15 | 20 | 17 | 14 |
| 3 | 2 | 1 | 2 | 4 | 4 | 1 | 6 | 3 | 3 | 1 |
| 4 | 3 | 1 | 2 | 0 | 1 | 3 | 3 | 1 | 2 | 2 |
| 5 | 7 | 14 | 11 | 18 | 18 | 28 | 40 | 43 | 53 | 56 |
Tabl. 19: Structure des dommages caus�s aux �pis avec intervention du pr�dateur Teretriosoma nigrescens (pourcentage d'�pis endommag�s sur un total de 300, � chaque date consid�r�e et dans les diverses cat�gories de dommages)
| Cat�gorie
de dommages |
Mois apr�s l'introduction du pr�dateur |
|||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | 50 | 56 | 56 | 54 | 50 | 53 | 46 | 43 | 39 | 39 |
| 2 | 35 | 29 | 25 | 25 | 30 | 26 | 23 | 27 | 28 | 27 |
| 3 | 3 | 2 | 3 | 3 | 3 | 1 | 2 | 3 | 3 | 1 |
| 4 | 4 | 4 | 2 | 2 | 0 | 1 | 3 | 1 | 2 | 2 |
| 5 | 8 | 9 | 14 | 17 | 19 | 18 | 28 | 28 | 27 | 31 |
4.2 Comparaison de l'efficacit� de Teretriosoma nigrescens au Togo et au Costa Rica
A la suite d'une comparaison entre des P. truncatus d'origine diverse, NISSEN (1989) a constat� de grandes diff�rences au niveau de la condition physique des populations. Dans le contexte d'une lutte biologique, la condition physique de la population de ravageurs serait susceptible d'exercer une nette influence sur l'efficacit� de la mise en oeuvre de l'insecte utile. L'essai d�crit ici visait � fournir une premi�re orientation en ce qui concerne l'efficacit� probable du pr�dateur au Togo par rapport � la r�gion d'origine commune au ravageur et � l'insecte utile. On a repris � cet effet les conditions d'essai expos�es dans les travaux de B�YE (1988), qui ont �t� r�alis�s au Costa Rica. La quasi-identit� des conditions d'essai permet une comparaison directe des r�sultats.
Mat�riels et M�thodes
20 P. truncatus adultes ont �t� install�s sur 200 g de ma�s en vrac, d'une esp�ce locale � grains blancs. Au bout de 7 jours, on a rajout� les T. nigrescens dans une proportion (P.t. contre T.n.) de "2 contre 1" et de "3,3 contre 1". Les essais ont dur� respectivement 50, 70 et 110 jours. Ils ont �t� effectu�s au moyen de r�cipients en verre de 1 I plac�s dans l'entr�e de la cage de toile m�tallique, dans laquelle r�gnaient une temp�rature de 19 � 38 �C et une humidit� relative variant de 25 � 99 %. L'interpr�tation est intervenue sur la base de la m�thode indiqu�e au chapitre 2.1.2.
R�sultats
La croissance d�mographique du ravageur a �t� fortement r�duite par la mise en oeuvre du pr�dateur, et cela ind�pendamment du rapport num�rique initial des individus (tabl. 20). Au cours de cet essai, le rapport �tait d�j� tellement �troit dans la variante "20 contre 6" que l'on en est arriv� � une r�duction maximale du nombre des ravageurs. La diff�rence entre le t�moin et les variantes impliquant le pr�dateur est all�e en augmentant au fur et � mesure que la p�riode d'essai s'avan�ait, ce qui est de toute �vidence d� � l'aptitude du pr�dateur � endiguer tr�s efficacement l'�volution d�mographique de P. truncatus.
Malgr� l'�troitesse du rapport num�rique entre les individus, T. nigrescens a �t� en mesure de se reproduire de mani�re tr�s satisfaisante (tabl. 20). La multiplication du pr�dateur �tait m�me l�g�rement meilleure dans la variante "20 contre 10". Au cours de l'essai, on a assist� dans les deux variantes � une r�duction suppl�mentaire du rapport num�rique proie-pr�dateur, qui est descendu jusqu'� 1,3 contre 1, bien que la valeur initiale ait fortement diverg� par rapport � celle rencontr�e dans les conditions naturelles, qui se situe aux alentours de 7,5 contre 1 (B�YE, 1988).
Tabl. 20: Evolution d�mographique de Prostephanus truncatus (P.t.) et de Teretriosoma nigrescens (T.n.) sur des grains de mais en fonction du rapport num�rique initial des individus
| P.t.:T.n. | P.
truncatus adultes apr�s...jours |
T.
nigrescens adultes apr�s ... jours |
||||
| 50 | 70 | 110 | 50 | 70 | 110 | |
| T�moin | 290 a | 710 a | 888 a | - | - | - |
| 20:10 | 26 b | 37 b | 48 b | 12 | 23 | 29 |
| 20:6 | 22 b | 38 b | 35 b | 8 | 16 | 26 |
En fonction de l'�volution d�mographique du ravageur, les param�tres de pertes accusaient �galement une baisse tr�s nette suite � la mise en oeuvre du pr�dateur (cf. tabl. 21). Dans le t�moin, les dommages avaient d�j� atteint 55,7 % au bout de 50 jours, c'est-�-dire plus du double du chiffre enregistr� dans les variantes de r�f�rence. Les dommages constat�s aux deux dates ult�rieures �taient eux aussi significativement r�duits � la suite de la mise en oeuvre du pr�dateur.
Tabl. 21: Dommages et pertes caus�s par Prostephanus truncatus (P.t,) sur des grains de mais en fonction du rapport num�rique initial des individus et compte tenu de la mise en oeuvre de Teretriosoma nigrescens (T.n.)
| P.t.:T.n. | Dommages
(en %) apr�s ... jours |
Pertes
(en %) apr�s ... jours |
||||
| 50 | 70 | 110 | 50 | 70 | 110 | |
| T�moin | 55,7 a | 83,9 a | 88,5 a | 14,7 a | 44,8 a | 63,3 a |
| 20:10 | 27,1 b | 47,4 b | 50,7 b | 4,9 b | 11,1 b | 17,2 b |
| 20:6 | 22,1 b | 48,8 b | 52,1 b | 4,4 b | 10,2 b | 15,2 b |
Au f�r et � mesure que l'essai se prolongeait, les pertes constat�es dans le t�moin pr�sentaient une forte augmentation, atteignant 63,3 % au bout de 110 jours. Si la progression s'est d�roul�e de fa�on analogue dans les variantes impliquant le pr�dateur, les pertes sont cependant demeur�es � un niveau largement inf�rieur. Au bout de 110 jours, les pertes enregistr�es dans ces variantes, qui avoisinaient 15 et 17 %, �taient respectivement inf�rieures de 76 et 73 % par rapport au t�moin.
P. truncatus s'est av�r� au cours de cet essai un antagoniste tr�s efficace contre P. truncatus. En comparaison des r�sultats des essais effectu�s au Costa Rica, on n'a pas constat� de diff�rence majeure du point de vue de l'efficacit�. Pour un rapport proie-pr�dateur de 2 contre l, le pr�dateur �tait en mesure de r�duire plus fortement la population du grand capucin du mais au Togo qu'au Costa Rica, alors que c'�tait l'inverse pour un rapport de 3,3 contre 1. On notera avec int�r�t l'�volution d�mographique plus favorable de P. truncatus dans les t�moins de l'essai d�crit ici, ce qui pourrait �tre l'indice de conditions environnementales �galement plus favorables et/ou d'une meilleure condition physique des col�opt�res (cf. NISSEN, 1989).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}