3.4 Spectre des h�tes de Nosema sp. et de Mattesia sp.
Table
des mati�res - ![]() Pr�c�dente
- Suivante
Pr�c�dente
- Suivante![]()
La transmissibilit� � d'autres esp�ces de col�opt�res des types de spores isol�s es sur P. truncatus permet de tirer certaines conclusions quant � la sp�cificit� des h�tes des deux esp�ces de protozoaires consid�r�es. Il est important de collecter des informations sur le spectre des h�tes de ces agents pathog�nes dans le mesure ou ces derniers pourraient �ventuellement infecter d'autres ravageurs des stocks et m�me, le cas �ch�ant bloquer leur �volution.
En vue des essais de transmission, on a s�lectionn�, outre P. truncatus, quatre esp�ces d'insectes �galement rencontr�es dans les greniers de stockage. Il s'agit d'une part d'un S. zeamais, ainsi que des ravageurs secondaires P. subdepressus, C. dimidiatus et T. castaneum. Deux bostryches, D. bifoveolatus et R. dominica ont Et! par ailleurs int�gr�s � cet essai du fait qu'il s'agit de ravageurs des stocks d'esp�ces voisines de P. truncatus et qu'ils nous ont paru � ce titre significatifs du point de vue de la transmission des protozoaires.
On trouvera au tableau 12 les r�sultats, d�taill�s pour chaque esp�ce de col�opt�re test�e, obtenus lors des essais d'inoculation des spores. Ce tableau renseigne �galement sur l'infection par Nosema sp. et Mattesia sp. pr�sent�e par chacun des stades de d�veloppement examin�s et pour chaque esp�ce d'insecte, ainsi que l'infection globale (totaux et pourcentages). Les spores de Nosema et de Mattesia ayant �t� inocul�es s�par�ment au bostryche D. bifoveolatus, les r�sultats d'infection d�taill�s figurent �galement � part. En ce qui concerne l'infection globale chez les col�opt�res de cette esp�ce, les r�sultats des deux inoculations ont �t� regroupes en un seul (cf. tabl. 12).
La profusion des r�sultats s�par�s figurent dans ce tableau permet de constater en premier lieu que toutes les esp�ces test�es ont �t� contamin�es par les protozoaires. A l'exception de T. castaneum, qui ne pr�sentait pas d'infection par Mattesia, les deux agents pathog�nes ont provoque une infection chez toutes les autres esp�ces de col�opt�res concern�es, ce qui exclut, aussi bien chez Nosema sp. que chez Mattesia sp., l'hypoth�se d'une sp�cificit� parasitaire au niveau du Grand Capucin du Ma�s.
Tabl. 12 : Infection cons�cutive � l'inoculation de spores de Nosema sp. (No. sp.) et de Mattesia sp. (Ma. sp.) � 6 esp�ces de ravageurs des stocks
| Esp�ce de coleopt�re |
Nombre des
individus examin�s |
Nombre des individus | |||||
| par les stades de d�veloppement |
somme | par | le total | ||||
| No. sp. | Ma. sp | Do* | somme | % | |||
| S. zeamais | 195 adultes | 14 | 9 | 2 | |||
| 7 pupes | 242 | 2 | 0 | 0 | 41 | 16,9 | |
| 40 larves | 10 | 3 | 1 | ||||
| P.subdepressus | 23 adultes | 19 | 0 | 2 | |||
| 8 larves | 31 | 5 | 0 | 3 | 29 | 93,5 | |
| C.dimidiatus | 14 adultes | 8 | 1 | 2 | |||
| 3 pupes | 100 | 2 | 0 | 1 | 87 | 87,0 | |
| 83 larves | 35 | 6 | 32 | ||||
| T.castaneum | 21 adultes | 2 | 0 | 0 | |||
| 37 larves | 58 | 20 | 0 | 0 | 22 | 37,9 | |
| R. dominica D.bifoveolatus |
15 adultes | 15 | 11 | 0 | 4 | 15 | 100 |
| 68 adultes | 10 | / | / | ||||
| 21 larves | 3 | / | / | ||||
| 176 | 41 | 23 3 | |||||
| 85 adultes | / | 27 | / | ||||
| 2 larves | / | 1 | / | ||||
* Do = Double infection
L'examen des divers r�sultats permet de constater que, compte tenu du nombre de sujets atteints d'une double infection, la microsporidie �tait en r�gle g�n�rale beaucoup plus fr�quemment repr�sent�e que la n�ogr�garine Dans les pr�parations � inoculer aux insectes, la concentration de spores de Nosema
Etait toutefois plus �lev�e que celle des spores de Mattesia (cf. chap 2.4.5), ce qui fait que les r�sultats in�gaux obtenus par chacun des agents pathog�nes sont vraisemblablement dus � cette diff�rence de densit� des spores au moment du traitement.
L'infection globale des diverses esp�ces de col�opt�res a donne des r�sultats tout a fait h�t�rog�nes. Alors que l on a relev� chez les ravageurs secondaires des taux d'infection allant de 38 % (sur T. castaneum) � 93,5 % (P. subdepressus), les individus de S. zeamais et de D. bifoveolatus n'�taient respectivement infestes par les protozoaires qu'� 17 et 23 %. On peut en d�duire que les ravageurs secondaires des stocks �taient de toute �vidence plus facilement contaminables par les protozoaires que les ravageurs des stocks aptes au forage. Notons cependant que l'on a constate une infection chez tous les repr�sentants du bostryche R. dominica, ce col�opt�re constituant en l'occurrence une exception.
Le tableau 12 permet par ailleurs de constater qu'il y avait � proportion, chez les ravageurs des stocks, davantage de larves infest�es que d'adultes, ce qui vaut aussi bien pour les ravageurs secondaires que pour les ravageurs primaires. S'agissant du bostryche R. dominica, il n'a pas �t� possible d'�tablir une comparaison entre les diff�rents stades de d�veloppement de cette esp�ce d'insectes vu que l'on a ici uniquement examin� des sujets adultes. Le fait que chez toutes les autres esp�ces de col�opt�res, ce soient les larves qui aient subi la plus forte infestation semble indiquer que ce stade (tait plus propice que celui de l'imago � une infection par des protozoaires, Les r�sultats obtenus sur P. truncatus ayant �t� analogues (cf. chap. 3.3.1 et 3.3.2), il ressort de l'essai de transmission que les protozoaires consid�r�s ont eu sur les h�tes secondaires des effets analogues � ceux observes sur l'h�te principal.
3.5 Transmission de Nosema sp. et de Mattesia sp. � Teretriosoma nigrescens
L'histeride T. nigrescens est un antagoniste de P. truncatus En tant que pr�dateur, il se nourrit des adultes et des larves du ravageur. La haute efficacit� qu'offre 7. nigrescens dans la lutte contre l'anobie fait de cet hist�rid� un antagoniste de tout premier plan du ravageur des stocks (HELBIG, 1993; P�SCHKO 1993). L'essaimage de l'insecte utile au Togo (B�YE et al , 1992) constituait le volet central du concept de lutte biologique contre le Grand Capucin du Ma�s.
A c�t� des nombreuses autres questions d'importance majeure sou lev�es par une telle d�marche, il �tait int�ressant de v�rifier si les deux esp�ces de protozoaires d�cel�es sur le ravageur �taient de nature � provoquer une infection chez T. nigrescens. Une affection �ventuelle par ces pathog�nes serait en effet susceptible d'entraver sensiblement l'�volution de l'hist�rid�, la mise en oeuvre future des protozoaires dans les greniers � odes paysans ayant par suite des cons�quences n�gatives pour le pr�dateur. Deux essais destines � �tudier le pr�disposition � l'infection de T. nigrescens ont �t� r�alis�s. Dans le cadre d'une premi�re installation exp�rimentale, on a plac� 30 T. nigrescens adultes dans un peuplement de P. truncatus fortement infestes par des protozoaires. Lors d'un second essai, on a donne en p�ture � des larves et adultes du pr�dateur quelques proies infect�es
Concernant les r�sultats, il faut noter que ni les larves, ni les adultes de T. nigrescens n'ont �t� infectes par les protozoaires. En outre, tous les P. truncatus places dans l'installation exp�rimentale �tant morts, on peut raisonnablement supposer que le pr�dateur s'�tait t effectivement nourri de ces proies.
Le diagnostic d'infection n�gatif �tabli au sujet de T. nigrescens corrobore les r�sultats de HUGER et P�SCHKO (communication interne), qui ont examin� quelque 1000 exemplaires de l'hist�rid� sans d�celer d'infection par les protozoaires. Il y a par cons�quent lieu de supposer que les spores de Nosema et de Mattesia trouv�es sur P. truncatus ne sont pas, en principe, en mesure d'infecter l'insecte utile.
3.6 Effets des insecticides associ�s � des spores de protozoaires en conteneur
3.6.1 Contagiosit� des spores apr�s
contamination par des insecticides (essai pr�liminaire)
3.6.2 Effets du pirimiphos-m�thyle associ� �
des spores sur la dynamique de population de Prostephanus
truncatus
En corr�lation avec les insecticides habituellement utilises dans les greniers � ma�s, l'application pratique de la pr�paration aux spores de protozoaires soul�ve deux questions. Il s'agit d'une part de mettre en lumi�re des interactions �ventuelles entre les spores de protozoaires et les ru�e es actives insecticides. L'application de spores dans un grenier � ma�s manquerait d'efficacit� si le pouvoir infectieux de ces spores �tait annihil� par une contamination due � des insecticides. Il �tait par ailleurs important de voir quels sont les effets des spores sur l'anoblie P. truncatus en cas d'application compl�mentaire d'une pr�paration chimique.
Afin de d�celer d'�ventuelles interactions entre les spores de protozoaires et certains insecticides, on a effectue un essai dit "pr�liminaire". Ce qualificatif veut dire que l'essai projet� �tait destine � fournir la base de l'examen ult�rieur visant � d�terminer les effets sur P. truncatus d'un traitement combine aux spores et aux insecticides. C'est dans le cadre d'un tel traitement combin� spores-insecticides que l'on a observe, en conteneur, la dynamique de populations du ravageur soumis � la pr�paration aux spores appliqu�e en association avec la mati�re active insecticide "pirimiphos-m�thyle".
3.6.1 Contagiosit� des spores apr�s contamination par des insecticides (essai pr�liminaire)
Apres avoir stocke les spores de la pr�paration mixte en contact, soit avec la mati�re active insecticide "pirimiphos-m�thyle" (Actellic), soit avec la mati�re active "deltam�thrine" (K-Othrine), on a teste sur des larves de P. truncatus le pouvoir infectieux des spores de Nosema et de Mattesia Les spores ayant �t� auparavant s�par�es de l'insecticide, chaque type de pr�paration aux spores a �t� inocul� � 40 larves de P. truncatus afin que l'on puisse en observer les effets infectieux sur ces larves. Une poudre aux spores conserv�e en �tat de non-contamination a servi de t�moin. Les quantit�s de spores appliqu�es Etaient identiques dans toutes les variantes (cf. chap. 2.4.6).
C'est l'inoculation de spores contamin�es � la deltam�thrine qui, avec 45 %, a donn� le taux d'infection global le plus �lev� chez les larves de P. truncatus utilis�es (fig. 24). Les spores contamin�es � l'ester d'acide phosphorique ont, quant � elles, provoque une infection chez 27,5 % des larves, tandis que 25 % seulement des larves de l'anoblie ont �t� infect�es � la suite d'un traitement � la poudre aux spores conserv�es i l'abri de la contamination par les insecticides. Le test Chi� a r�v�l� que ces taux d'infection ne pr�sentaient entre eux aucune diff�rence significative.
Comme le montre par ailleurs la figure 24, les deux types de protozoaires se sont manifestes dans toutes les variantes, Compte tenu des sujets atteints d'une double infection, les larves de l'anobie �taient plus fr�quemment infect�es par Nosema sp. que par les esp�ces Mattesia. Ce r�sultat est conforme du fait que la pr�paration appliqu�e renfermait une quantit� proportionnellement sup�rieure de spores de Nosema (cf. Chap. 2.4.6). On ne peut donc pas en conclure que les deux types de spores diff�rent au niveau du pouvoir infectieux apr�s contamination par les insecticides consid�r�s.
L'infection subie par les larves permet de conclure en r�sume que la contamination par les mati�res actives insecticides n'a pas amoindri la contagiosit� des deux types de spores. Le taux d'infection relativement �lev� des deux agents pathog�nes constat� sur les larves � la suite du traitement aux spores contamin�es par de la deltam�thrine permet m�me de supposer que le stockage commun avec l'insecticide "K-Othrine" stimule le pouvoir infectieux des spores.
L'essai d�crit dans la suite avait pour but de d�terminer les effets, sur la population de P. truncatus, de traitements aux spores de protozoaires associ�es au pirimiphos-m�thyle, une mati�re active insecticide. Nous allons bri�vement indiquer ici les raisons qui ont pr�side du choix d'une liaison ester d'acide phosphorique pour une mise en oeuvre combin�e � des spores de protozoaires.
On sait par les ouvrages sp�cialis�s que la mati�re active "pirimiphos-m�thyle" n'est pas en mesure de paralyser durablement le d�veloppement de P. truncatus (GOLOB et a 1, 1985). Cette efficacit� insuffisante de l'ester d'acide phosphorique nous a conduits � ver fier si l'association du principe actif a des spores de protozoaires �tait de nature � en renforcer l'efficacit�. L'id�e de d�part �tait que la contamination de l'insecte par les protozoaires pouvait �ventuellement le sensibiliser davantage au principe actif chimique.
On a pris en l'ocurrence des m�les et femelles sains du ravageur des stocks, que l'on a pinc�s dans des r�cipients � raison de 10 sujets par conteneur. Ces animaux ont re�u respectivement un traitement unique � la pr�paration aux spores, au principe actif "pirimiphos-m�thyle, ou encore ils ont �t� trait�s du moyen de pr�parations combin�es de poudre aux spores et de pirimiphos-m�thyle. Les doses employ�es lors de l'application de l'insecticide seul et des traitements combin�s �talent respectivement de 0.01 et de 0.005 mg de pirimiphos-m�thyle par g de substrat (cf. tabl. 3).
Il a �t� proc�d� � r�p�titions par variante. A l'issue d'une p�riode de 4, 8 et 12 semaines, les individus ont �t� d�nombr�s dans chacune des variantes. Dans les peuplements traites aux spores ou aux pr�parations combin�es, on a pr�lev� � l'issue de chacune des p�riodes indiqu�es un �chantillonnage de 50 animaux par stade de d�veloppement et proc�d� sur eux � une recherche d'infection par les protozoaires.
Si l'on compare le d�veloppement de P. truncatus sous l'effet des divers traitements, on constate que ce sont les populations non trait�es qui ont connu le d�veloppement la plus favorable (fig. 25, t�moin). On a pu observer pendant toute la dur�e de l'essai une croissance de ces populations, qui comptaient au bout de 12 semaines 2390 animaux en moyenne. Les populations de ravageurs trait�es � la pr�paration aux spores ont connu un d�veloppement plus mauvaise que le t�moin (fig. 25, spores). Avec quelque 1500 sujets au bout de 12 semaines, les animaux �taient en l occurrence nettement moins nombreux qu'au sein de la population non trait�e.
Sous l'effet du pirimiphos-m�thyle, le nombre de sujets � se d�velopper a �t� dans tous les cas inf�rieur � ce qu'il a �t� sur le t�moin et � la suite de l'application de spores. Si le traitement � 0,005 mg de pirimiphos-m�thyle par g de substrat (fig. 25, Actellic 1) a permis d'observer pendant toute la dur�e de l'essai une augmentation du nombre d'individus, on a constate en revanche apr�s 12 semaines une r�duction moyenne des populations � 612 animaux seulement. Les populations trait�es � 0,01 mg de pirimiphos-m�thyle par g de substrat se sont reproduites dans l'ensemble encore plus mal (fig. 25, Actellic 2), puisqu'elles ne comptaient plus en moyenne que 114 individus au bout de 12 semaines.
Dans les conteneurs traites aux pr�parations combin�es, la dynamique de la population s'est maintenue pendant toute la dur�e de l'essai � un niveau sup�rieur � ce qu'elle �tait chez les P. truncatus traites � l'Actellic (cf. fig. 25). Ayant d�j� atteint une densit� relativement �lev�e au bout de 4 semaines d'essai, les populations ont �t� en mesure de continuer � se multiplier �galement apr�s 8 semaines' et cela pour les deux traitements combines. Au bout de 12 semaines, on a d�nombr� en moyenne dans la variante trait�e aux spores additionn�es de 0,005 mg de pirimiphos-m�thyle, par g de substrat (traitement combin� 1) 500 animaux. On a constat� ici un l�ger recul du nombre de sujets par rapport � ce qu'il �tait au bout de 8 semaines. Les populations trait�es � 0,01 mg de principe actif par g de substrat additionn� de spores (traitement combin� 2) comptaient au bout de 12 semaines 367 individus, soit � peu pr�s le m�me nombre qu'au bout de 8 semaines.
Le taux de mortalit� des populations d'anobies ayant subi les divers traitements a �t� �tabli � chaque fois pour tous les individus d'un m�me conteneur. Le tableau 13 indique les taux de mortalit� moyens par variante � chacune des dates d�valuation. Ce sont les populations trait�es � l'Actellic hautement concentr� (Actellic 2) qui pr�sentaient le taux de mortalit� le plus �lev�. Au bout de 8 semaines, on d�nombrait en effet ici 70,1 % d� sujets morts et 65 4 % au bout de 12 semaines. Dans toutes les autres variantes en revanche, le nombre de col�opt�res morts atteignait tout au plus 33,6 %, ce qui se situe nettement en de�� des taux de mortalit� des populations trait�es � l'Actellic 2. On notera en l'occurrence la mentalit� initiale �lev�e observ�e au sein des populations de ravageurs trait�es uniquement a l'insecticide. Au bout de quatre semaines, le taux de mortalit� s'�tablissait respectivement � 30 % (Actellic 1), et � 36 % (Actellic 2), alors que les populations des autres variantes accusaient des taux de mortalit� allant de 1 � 6 % seulement.
Tabl 13: Mortalit� chez Prostephanus truncatus � la suite de divers traitements � la pr�paration aux pores et � l'Actellic apr�s 4,8 et 12 semaines a essai
| Variantes | Mortalit� (%) |
||
| 4 | 8 | 12 | |
| temoin | 3,3 | 10,5 | 27,1 |
| spores | 1,3 | 21,3 | 33,6 |
| Actellic 1 | 29,9 | 29 ,8 | 21,0 |
| Actellic 2 | 36,3 | 70,1 | 65,4 |
| traitement | 3,4 | 22,6 | 33,5 |
| combin�s 1 | |||
| traitement | 6,2 | 11,3 | 17,6 |
| combin�s 2 | |||
* variantes: pour les abr�viations, se reporter � la fig. 25.
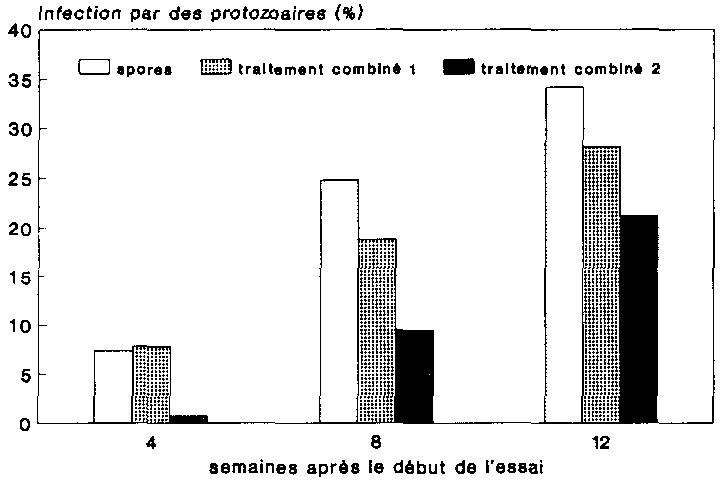
On trouvera sur la figure 26 les r�sultats de l'infection par les protozoaires chez les populations trait�es aux spores (spores, traitements combin�s 1 et 2). En ce qui concerne les taux d'infection, il s'agit de l'infection globale moyenne pour tous les stades de d�veloppement � une date donn�e, 50 sujets ayant �t� examines � chaque fois.
On a observe pour toutes les variantes une augmentation des taux d'infection � chaque date d'�valuation. A la diff�rence des taux d'infection obtenus � la suite du traitement simple aux spores, les traitements combines se sont traduits par des taux d'infection moindres au niveau des individus. On notera ici avec int�r�t que le traitement combin� comportant la concentration de pirimiphos-m�thyle plus faible (traitement combin� 1) a provoque au sein des populations un taux d'infection proportionnellement sup�rieur � celui r�sultant du traitement qui faisait intervenir une concentration de mati�re active plus �lev�e (traitement combin� 2).
Les diff�rences constat�es d'une variante a l'autre indiquent clairement que l'application de spores seules a permis aux protozoaires de se propager plus facilement dans la population de l'H�te que lorsqu'elles ont �t� appliqu�es en association avec un traitement a l'Actellic. Il semble que la concentration de mati�re active insecticide introduite dans le conteneur ne soit pas sans influence sur la propagation de l'agent pathog�ne, puisque aussi bien les traitements hautement concentres ont �t� suivis d'un taux d'infection � proportion plus faible que ceux qui contenaient une moindre concentration de mati�re active.
C'est � l'issue de 12 semaines d'essai que l'on a d�termin� chez les populations trait�es aux spores le taux d'infection le plus �lev�, c'est-�-dire 34 %. Les autres taux d'infections �tant demeures inf�rieurs � 30 %, l'infection s'est donc av�r�e faible dans l'ensemble. Il n'est pas possible, du moins pour l'instant, de tirer des conclusions quant � ce bas niveau d'infection vu que les doses de spores inocul�es �taient relativement �lev�es. Il est toutefois concevable que la pr�paration inocul�e n'ait �t� que faiblement infectieuse.
En r�sum�, on peut dire que les traitements aux spores de protozoaires et ceux � la mati�re active pirimiphos-m�thyle ont eu des effets r�ducteurs sur la dynamique des populations de P. truncatus. L'application des variantes aux insecticides s'est traduite par des taux de mortalit� relativement �lev�s qui n'ont �t� constat�s ni pour les traitements combines, ni sous le seul effet des spores. Compar�es aux populations trait�es � l'Actellic, celles qui avaient �t� trait�es au moyen des pr�parations combin�es ont produit dans l'ensemble davantage de sujets. Ces r�sultats permettent de conclure que le m�lange de poudre insecticide et de poudre aux spores donne vraisemblablement naissance � des interactions antagoniques entre la mati�re active insecticide et les spores de protozoaires. Cette hypoth�se expliquerait d'une part le baisse d'efficacit� de l'insecticide contre le ravageur, et d'autre part les faibles taux d'infection sur P. truncatus dans les variantes � pr�parations combin�es.
Ajoutons pour terminer qu'il n'a pas �t� possible de tirer de conclusions quant aux effets de l'ester d'acide phosphorique sur des P. truncatus infectes par des protozoaires en raison du faible niveau d'infection constat� dans l'ensemble.
3.7 Mise en �vidence de spores de protozoaires dans des produits r�siduels du stockage du ma�s
Du point de vue de la mise en oeuvre pratique des spores de protozoaires il serait souhaitable, et cela pour des raisons �videntes de r�duction des co�ts, que les paysans se chargent eux-m�mes de confectionner la pr�paration aux spores de protozoaires. Nous allons examiner dans la suite les possibilit�s dont dispose l'utilisateur pour fabriquer cette pr�paration de mani�re autonome.
Au cours du processus d'extraction des grains cons�cutif du stockage du ma�s en �pis tel qu'il est traditionnellement pratique dans le sud du Togo, il se forme des r�sidus n'ayant aucune valeur nutritive. Il s'agit en l'occurrence de spathes, de rachis, de brisures de grains, d'ivraie, et, en cas d'infestation de l'entrep�t par des ravageurs' d'insectes parasites et de la poussi�re de forage qu'ils produisent. Afin d'�tudier les possibilit�s de fabrication d'une poudre aux spores � partir des insectes existants, on a collecte les r�sidus d'un stockage de ma�s, que l'on a ensuite examin�s � la recherche de spores �ventuelles.
Durant une p�riode de cinq mois, on a pr�lev� chaque mois dans un grenier � ma�s 50 �pis Les r�sidus provenant de l'extraction des grains ont �t� stockes dans un fut pratiquement herm�tique. Apres avoir op�r� le dernier pr�l�vement d'�pis dans le grenier, les d�chets conserv�s ont �t� tri�s, et les �l�ments grossiers, � savoir les spathes, les axes des �pis et les brisures de grains, ont �t� s�par�s des r�sidus fins. A partir du m�lange r�siduel, constitu� de poussi�re de farine, de col�opt�res et d'ivraie, on a pr�par� une poudre fine, dont on a pr�lev� trois �chantillons de 20 mg chacun. Ces �chantillons de poudre ont �t� ensuite examin�s selon la m�thode expos�e plus haut afin de d�terminer la densit� des spores (cf. chap. 2.3.5).
L'analyse des �chantillons de farine a r�v�l� une concentration moyenne de 7,5x104 de spores de Mattesia et de 5x105 de spores de Nosema dans I ml d'eau distill�e. Compar�es � celles des pr�parations utilis�es lors des essais en conteneurs, ces valeurs �taient inf�rieures de 1 � 2 puissances de dix. On a donc mis en �vidence dans la poudre obtenue � partir des r�sidus de stockage du ma�s une concentration de spores tr�s �lev�e. Les spores de protozoaires d�tect�es dans la farine proviennent vraisemblablement des cadavres d'insectes contenus dans le m�lange pulv�ris�.
3.8 Application de la pr�paration aux spores de protozoaires dans des conditions semi-pratiques
3.8.1 Mise en oeuvre de la pr�paration aux
spores dans un grenier � ma�s traditionnel-essai I (1990) �
Cacaveli
3.8.2 Mise en oeuvre de la pr�paration aux
spores dans un grenier � ma�s traditionnel - essai II (1990/91)
� Davi�
Apr�s examen des r�sultats des essais r�alis�s en conteneurs, nous allons d�crire dans la suite ceux qui avaient pour but de tester, dans des conditions proches de la r�alit�, les effets de la pr�paration aux spores. Deux essais similaires de stockage, r�alis�s selon les m�thodes traditionnelles. de stockage du ma�s en usage dans cette r�gion, se sont d�roul�s dans le sud du Togo. Dans le cadre de l'essai de stockage I, le ma�s a �t� entrepose de f�vrier � juillet 1990 (pendant la "petite saison de stockage"), tandis que l'essai II avait lieu d'ao�t 1990 � janvier 1991 (durant la "grande saison").
{kind=link}
{kind=link}
{kind=link}